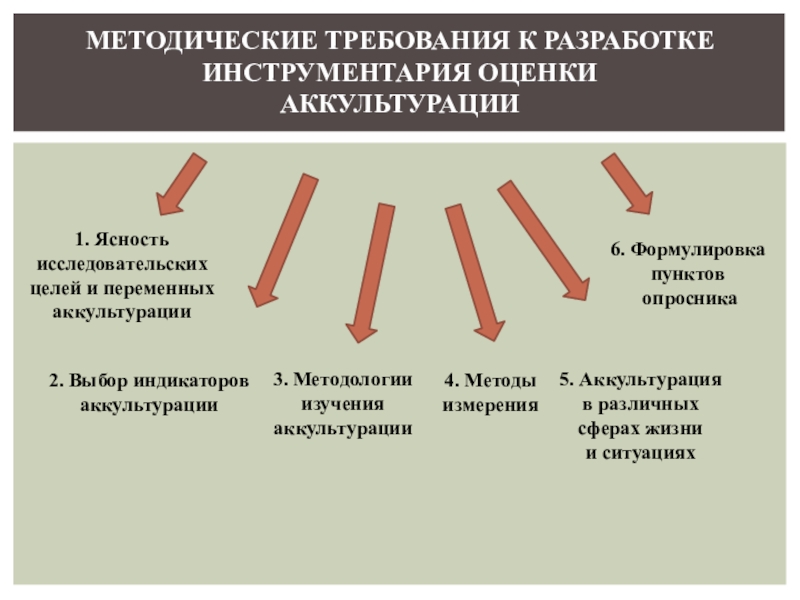
Слайд 1Лекция 2
Морфология и функции поверхностного аппарата клетки
1. Общая морфология клетки
2.
Строение и функции биологических мембран
3. Надмембранный комплекс, его структурно-функциональная организация
4.
Межклеточные контакты, их строение
5. Субмембранный комплекс – цитоскелет
6. Транспорт веществ через мембраны:
а) пассивный
б) активный
7. Участие биомембран в передаче информации
Слайд 2Поверхностный аппарат клетки
состоит из:
- плазмалеммы (1),
надмембраннного (2)
субмембранного
(3)
комплексов
Слайд 3 Поверхностный аппарат клетки
состоит из:
- плазмалеммы (1),
надмембраннного (2)
субмембранного (3)
комплексов
1. Плазмалемма образована:
белками
(~ 60%)
липидами (~ 40%)
← гидрофильная и
← гидрофобная части
- фосфолипиды, гликолипиды, стеролы
белки: интегральные (а),
полуинтегральные (б),
периферические (в)
2. Надмембранный комплекс - выполнен гликолипидами и гликопептидами
- функция. №1 – рецепция
- в животной клетке – гликокаликс,
- в растительной клетке гликокаликс + клеточная стенка,
- грибы, насекомые – хитин
3. Субмембранный слой - скопление микротрубочек и микрофиламентов цитоскелета под плазмалеммой.
Слайд 4II. Поверхностный аппарат клетки
состоит из:
- плазмалеммы (1),
надмембраннного (2)
субмембранного (3)
комплексов
Функции поверхностного аппарата клетки:
- барьерная,
-
обменно-транспортная,
- рецепторная,
- метаболическая (ферментативная),
- контактная (в многоклеточном организме),
- опорно-сократительная,
- генетическая индивидуальность или гистосовместимость …
Слайд 5Классификация и характеристика мембранных липидов
Липиды
(30 -70%)
фосфолипиды
гликолипиды
стероиды
сфинголипиды
глицерофосфоли-пиды
цереброзиды
сульфатиды
ганглиозиды
Холестерин (животные)
Ситотерин, стигмастерин
(растения)
Эргостерин (грибы)
Слайд 6Общий принцип строения мембранных липидов
Слайд 8Мембранные белки
Мембранные белки классифицируются по их положению в мембране:
поверхностные;
интегральные ;
полуинтегральные.
Слайд 9По функциональной роли выделяют 4 группы белков:
Структурные белки – придают
клетке и органеллам определенную форму; придают мембране механические свойства (эластичность);
обеспечивают связь мембраны с цитоскелетом.
Транспортные белки.- создают устойчивые транспортные потоки определенных веществ через мембрану, обеспечивают возникновение трансмембранного потенциала во всех клетках.
Белки, обеспечивающие межклеточные взаимодействия. Их подразделяют на два класса: 1. адгезивные белки; 2. белки, участвующие в образовании специализированных межклеточных контактов.
Белки, участвующие в клеточной сигнализации. Выделяют 3 функциональных вида:1. рецепторные белки; 2 белки трансмиттерного устройства, передающие сигнал через мембрану; 3. ферменты, одни из которых на внешней поверхности инактивируют сигнальные вещества, а другие на внутренней поверхности реагируют на сигнал образованием внутриклеточного медиатора.
Слайд 10Жидкостно-мозаичная модель строения биологических мембран
(Nikolson, Singer, 1972)
Слайд 11Надмембранный комплекс. Циторецепторы.
Поверхностные рецепторы имеют три домена: 1. внеклеточный эктодомен,
2. трансмембранный домен, 3 цитоплазматический домен. Эктодомен содержит участок связывания
сигнальной молекулы или лиганда. Обычно это самая большая часть белкового рецептора. Трансмембранные домены рецепторов можно разделить на две группы: первая – семейство рецепторов, пронизывающих мембрану семь раз – их называют «серпантинными» рецепторами (например, β2-адренергический рецептор, родопсин); второе – семейство рецепторов, пронизывающих мембрану один раз – это монотопные рецепторы, они обладают собственным ферментным участком, расположенным в цитоплазме. Цитоплазматический домен содержит действующие ферментативные центры, его каталитическая способность обусловлена аминокислотной последовательностью.
Слайд 12внутриклеточные
поверхностные
рецепторы
Каталитические рецепторы
Рецепторы, связанные с ионными каналами
Рецепторы, связанные с G-белками
Рецепторы, связывающие
молекулы внеклеточного матрикса с цитоскелетом
Цитоплазматические рецепторы
Внутриядерные рецепторы
Слайд 13Молекулы межклеточной адгезии
1. Кадгерины - трансмембранные гликопротениы, использующие для адгезии
ионы кальция. Отвечают за организацию цитоскелета, взаимодействие клеток с другими
клетками.
Слайд 142. Интегрины - представляют собой мембранные рецепторы для белковых молекул
внеклеточного матрикса – фибронектина, ламинина и др. Связывают внеклеточный матрикс
с цитоскелетом при помощи внутриклеточных белков талина, винкулина, α-актинина. Функционируют как клеточно-внеклеточные, так и межклеточные адгезионные молекулы.
Слайд 153. Селектины - обеспечивают прилипание лейкоцитов к эндотелию сосудов и
тем самым – лейкоцитарно-эндотелиальные взаимодействия, миграцию лейкоцитов через стенки сосудов
в ткани.
Слайд 16Семейство иммуноглобулинов - эти молекулы играют важную роль в иммунном
ответе, а также в эмбриогенезе, заживлении ран и др.
Слайд 17Гоминговые молекулы - обеспечивают взаимодействие лимфоцитов с эндотелием, их миграцию
и заселение специфических зон иммунокомпетентных органов.
Слайд 18Межклеточные контакты
Межклеточные контакты
Простые
Сложные
Запирающие
Сцепляющие
Коммуникационные
Плотный контакт
Адгезивный поясок
Десмосомы
Щелевой контакт
Синапс
Плазмодесмы
Интердигитация
Простое
соединение плазмалемм
Слайд 19Простые контакты
Простое межклеточное соединение представляет собой сближение плазмолемм соседних клеток.
Гликопротеиды соседних клеток при образовании простого контакта «узнают» клетки одного
типа. Большая часть поверхности контактирующих клеток эпителия связана простыми контактами. Причем существуют специальные белки-рецепторы (Е-кадгерины), участвующие в образовании контактов только между эпителиальными клетками.
Слайд 20Плотный (запирающий) контакт
К запирающим относится плотный контакт. В этом соединении
принимают участие специальные интегральные белки, расположенные на поверхности соседних клеток,
образующие подобие ячеистой сети. Эта ячеистая сеть окружает в виде пояска весь периметр клетки, соединяясь такой же сетью на поверхности соседних клеток. Эта область не проницаема для макромолекул, и следовательно, она запирает, отграничивает межклеточные щели от внешней среды. Этот вид соединения характерен для клеток однослойных эпителиев и эндотелия.
Слайд 21Адгезивный поясок
Адгезивный (сцепляющий) поясок – парное образование в виде ленты,
опоясывающей апикальную часть клетки однослойных эпителиев. Здесь клетки связаны друг
с другом интегральными гликопротеидами к которым со стороны цитоплазмы и той и другой клетки примыкает слой примембранных белков, включающих характерный белок винулин. К этому слою подходит и связывается с ним пучок актиновых микрофиламентов.
Слайд 26Субмембранный комплекс - цитоскелет
Слайд 27Мембранным транспортом называют любой переход атомов, ионов, молекул веществ через
мембрану из среды в клетку или в обратном направлении независимо
от путей, сил и механизмов переноса.
В зависимости от характера связи транспорта данного иона металла или молекулы конкретного вещества от переноса других ионов или молекул веществ выделяют:
-унипорт ‑ транспорт ионов или молекул через мембрану независимо от транспорта других соединений, например, молекул газов, воды;
-симпорт ‑ одновременный и однонаправленный перенос ионов или молекул двух различных веществ, например, перенос ионов Na+ и глюкозы через мембрану клеток эпителия тонкой кишки;
-антипорт ‑ одновременный транспорт ионов или молекул веществ через мембрану в противоположных направлениях.
Слайд 28Каналы – образования белковой природы (чаще всего крупные молекулы белка),
имеющие центральную полость для прохождения преимущественно ионов одного вида (Na+,K+,Cа2+,H+)
и механизмов, обеспечивающих их открытие и закрытие.
Слайд 29Переносчики ‑ молекулы веществ, осуществляющие транспорт ионов или молекул в
результате временного взаимодействия их с веществом. В роли переносчиков выступают
ионофоры, сиаловые кислоты, белки.
Слайд 30Насосы – мембранные АТФазы, специализирующиеся на противоградиентном переносе ионов Na+,
K+, Ca2+, H+ за счет энергии гидролиза АТФ.
Слайд 31Пассивный транспорт идет по градиенту концентрации веществ, без затраты энергии
(АТФ)
О2, СО2, N2,
бензол,
мочевина,
глицерин,
Н2О
"Понг"
Пассивный транспорт
Движущими силами пассивного переноса веществ
через мембрану служат градиенты:
концентрационный – для нейтральных молекул;
электрохимический – для ионов;
осмотический и градиент гидростатического давления – для воды;
градиент парциальных давлений для газов.
Слайд 32Катионные каналы
Калиевый канал
Натриевый канал
Слайд 33Никотиновый ацетилхолиновый рецептор
Слайд 34Активный транспорт можно разделить на 2 вида:
ак.тр. низкомолекулярных
соединений,
- ак.тр. высокомолекулярных соединений.
Активный транспорт низкомолекулярных соединений. Пример: (Na+K+) –
насос. Антипорт.
Активный транспорт
Слайд 35Принцип работы натрий-калиевой АТФазы
Слайд 36Вторично-активный транспорт
Na+–Ca2+‑обменный переносчик переносит из клетки Ca2+ в обмен на
2 Na+, используя энергию электрохимического градиента Na+, который генерируется на
мембране благодаря активности NаК – АТФаза
Натрий-кальциевый обменный переносчик
Слайд 37Активный транспорт высокомолекулярных соединений.
в → клетку – эндоцитоз:
жидких в-в → пиноцитоз
твердых в-в → фагоцитоз
из ← клетки – экзоцитоз
Пример: Микропиноцитоз
а. рецепция,
б. присоединение клотриновых белков (окаймление),
в. впячивание – инвагинация плазмолеммы с образованием окаймленной ямки,
г. обособление эндоцитозного пузырька (окаймленный пузырек, эндосома, пиносома, пиноцитозный пузырек, прелизосома),
сброс каймы.
Слайд 38Рекреция - перенос твердых веществ через клетку
Слайд 39Биомембраны – важный компонент систем регуляции процессов, протекающих в организме,
т.к. они содержат и высокоспециализированные рецепторы, принимающие сигналы и эффекторы,
регулирующие и преобразующие поступающие сигналы.
Процесс переработки информации, поступающей как снаружи, так и изнутри организма, подразделяют на три стадии: 1) получение информации рецепторами,
2) передача информации в центр переработки,
3) передача обратного стимула к устройствам, способным сформировать ответ – эффекторам.
Слайд 40Передачу сигналов от клетки к клетке осуществляют первичные мессенджеры.
Гормоны
Нейромедиаторы
Гистогормоны
Основные механизмы действия первичных мессенджеров:
Вещество взаимодействует с рецептором
плазмалеммы, что вызывает химическую модификацию белков (фосфорилирование, дефосфорилирование)
Вещество взаимодействует с рецептором плазмалеммы, который одновременно является ионным каналом, открывающимся при связывании регулятора
Вещество проникает внутрь клетки мишени, связывается с цитоплазматическим белком рецептором, и выступает как транскрипционный фактор, влияя на экспрессию генов.
Слайд 41Взаимодействие стероидных гормонов с клеткой (гуморальная регуляция)
Слайд 42Взаимодействие белково-пептидных гормонов с клетками
(гуморальная регуляция)
Слайд 44Передача сигналов с помощью нейромедиаторов
Слайд 45Основные механизмы передачи внутриклеточных сигналов
Фосфорилирование
Дефосфорилирование
Изменяет конформацию белков
Активирует ферменты, проявляющие
киназную активность
Создает в молекулах белков стыковочные участки
Запускает механизмы инактивации сигнала:
Поглощение
сигнального лиганд-рецепторного комплекса путем эндоцитоза
Десенсибилизация рецептора
Разрушение эффекторной молекулы
Слайд 46Передача сигналов с помощью G-белков и вторичных мессенджеров
Аденилатциклазный путь
Слайд 48Передача сигнала в клетку под влиянием адреналина

, надмембраннного (2) субмембранного (3) комплексов")
, надмембраннного (2) субмембранного (3)")
, надмембраннного (2) субмембранного (3)")
 фосфолипидыгликолипидыстероидысфинголипидыглицерофосфоли-пидыцереброзидысульфатидыганглиозидыХолестерин (животные)Ситотерин, стигмастерин (растения)Эргостерин (грибы)")




")









 контактК запирающим относится плотный контакт. В этом соединении принимают")
 поясок – парное образование в виде ленты, опоясывающей")

")




, имеющие")


О2,")









")
")