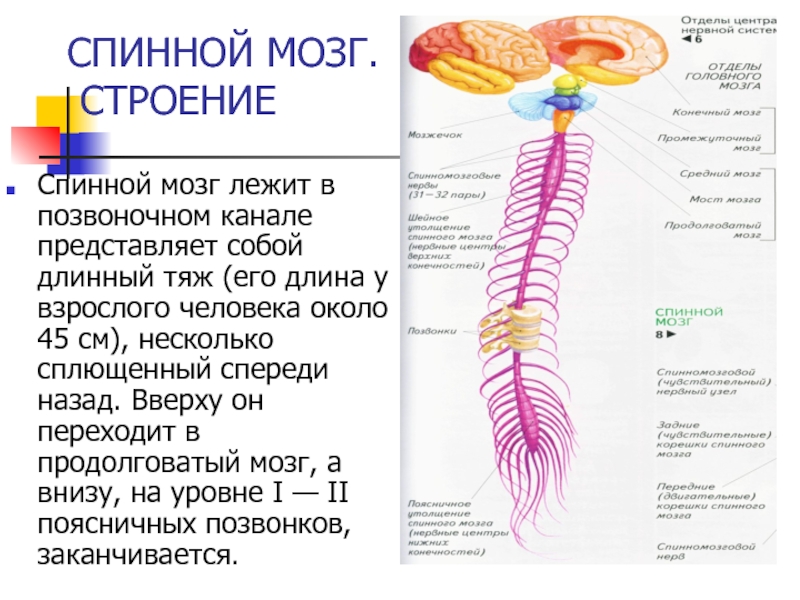
царствами – архебактерии и эубактерии
Домен – консервативная последова-тельность аминокислот, присутствую-щая
в нескольких (обычно во многих) белковых молекулах у разных организ-мов. Большинство доменов характери-зуются строго определенной функцией и представляют собой функциональ-ные блоки белковых молекул.
В геноме эукариот белки, ответствен-ные за операции с геномом (реплика-ция, транскрипция, трансляция) и белки, осуществляющие операции с мембранами – от архей, белки, белки основного обмена – от эубактерий.
существует гипотеза, что первичный прокариотический безъядерный организм, образован путем слияния архебактерии с эубактерией, причем базовый энергетический метаболизм этого организма имел эубактериальную природу (гликолиз, брожение)
Количественное соотношение общих и уникальных белковых доменов у архей, бактерий и эвкариот. Площади фигур примерно пропорциональны числу доменов (из А.В.Марков, А.М.Куликов, 2004).







Росомаха питается")





 и различия")