Слайд 1Метаболизм
Переход от катаболизма к анаболизму
I. Синтез липидов
Слайд 2осуществляется из углеводов, поступающих в избыточном количестве
участвуют некоторые аминокислоты
Накоплению жиров
способствует и избыток пищи
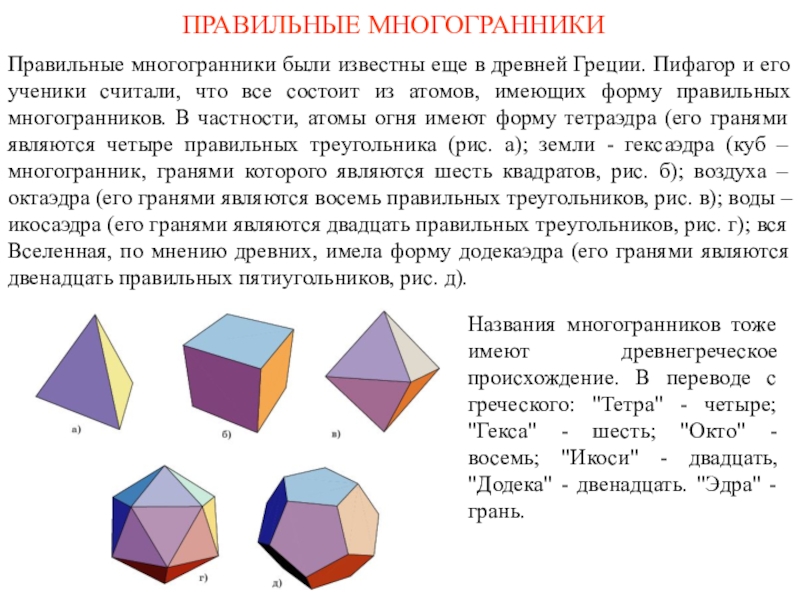
Жир – наиболее компактная форма запасания энергии
организмом
Синтез жиров
Слайд 4Синтез ЖК в тканях протекает в цитоплазме клеток и резко
отличается от процесса их окисления
Строительный блок – ацетил-КоА
Для синтеза ЖК
необходимо наличие в цитоплазме СО2 и НСО3–
Слайд 5Синтез ЖК начинается с действия полифункционального фермента ацетил-КоА-карбоксилазы, содержащего
биотин (витамин
Н)
домен биотинкарбоксилазы
биотинкарбоксилпроводящий домен
домен транскарбоксилазы
Данный фермент ускоряет синтез малонил-КоА
Слайд 61)
2) Карбоксибиотин + СН3СОSКоА →
→ НООС–СН2–СОSКоА + биотин
Малонил-КоА
Слайд 7Последующие стадии синтеза ЖК осуществляются при посредстве второго полифункционального фермента
– синтетазы высших жирных кислот, содержащего
3 домена и 8 субдоменов,
в частности, ацилпроводящий домен (АПД), функция которого – передвижение ацильной группы от одного субдомена к другому в соответствии с химизмом
Слайд 8Последовательность реакций синтеза ВЖК
Слайд 9Домен I (элонгация цепи)
СН3–СОSКоА + НS–АПД
НSКоА +
СН3–СОSАПД
ацетил-АПД
субдомен АПД-ацетилтрансферазы
НООС–СН2–СОSКоА + НS–АПД
НООС–СН2–СОS–АПД
+ НSКоА
малонил-АПД
субдомен АПД-малонилтрансферазы
СН3–СОSАПД + НООС–СН2–СОS–АПД
СН3–СО–СН2–СОSАПД + НS–АПД + СО2
ацето-ацетил-АПД
субдомен 3-кетоацилсинтетазы (конденсирующий субдомен)
Слайд 10Домен II (восстановление)
СН3–СО–СН2 –СОSАПД + НАДФН2
СН3–СН(ОН)–СН2–СОSАПД
+ НАДФ
-гидроксибутирил-АПД
субдомен -кетоацилредуктазы
СН3–СН(ОН)–СН2–СОSАПД
Н2О + СН3–СН=СН–СОSАПД
кротонил-АПД
субдомен -оксиацилдегидратазы
СН3–СН=СН–СОSАПД + НАДФН2
СН3–СН2–СН2–СОSАПД
бутирил-АПД
субдомен еноилредуктазы
Далее цикл реакций повторяется
В результате прохождения одного цикла углеродная цепь кислоты удлиняется на 2 атома С
Слайд 11Домен III (тиолиз, завершение синтеза)
СН3–(СН2)14–СОS–АПД + Н2О
СН3–(СН2)14–СООН + НS–АПД
пальмитил-АПД
Тиоэстераза
Слайд 12Суммарное уравнение синтеза пальмитиновой кислоты
8СН3–СОSКоА + 7АТФ + 14
НАДФН2
СН3–(СН2)14–СООН + 14 НАДФ +
+ 8 НSКоА +
7 АДФ + 7 Н3РО4 + 6Н2О
Слайд 13Схема синтеза ЖК
Ацетил-КоА
(n+1) моль
1 моль
ацетил-КоА
n моль
малонил-КоА
ВЖК
С2(n+1)
Слайд 14Синтез ненасыщенных ЖК
Пальмитолеиновая и олеиновая кислоты – из пальмитиновой
и стеариновой под действием специфической монооксигеназной ферментной системы, локализованной в
ЭПС, при участии цит b5 (в разных тканях, в том числе – в печени)
СН3–(СН2)14–СОSКоА + НАДФН2 (или НАДН2) + О2
пальмитил-КоА
СН3–(СН2)5–СН=СН–(СН2)7–СОSКоА + НАДФ (НАД) + 2Н2О
пальмитоолеил-КоА
Линолевая и линоленовая кислоты не синтезируются тканями животных и человека и должны поступать с пищей (?), поэтому их называют незаменимыми (витамин F)
Слайд 15Биосинтез триглицеридов
Три различных пути
Слайд 16В слизистой оболочке кишечника
(см. выше)
1. Из -моноглицеридов
Слайд 172. Из глицерина и жирных кислот
Путем реакций трансацилирования
В печени,
слизистой оболочке кишечника, жировой ткани и т. п.
На мембранах ЭПС
клеток
Слайд 181). Образование -фосфоглицерина
2 пути:
фосфорилирование глицерина (в почках и у
микроорганизмов)
Глицеринкиназа
Слайд 191). Образование -фосфоглицерина
2 пути:
восстановление ДОАФ (в мышцах, в слизистой
кишечника и жировой ткани)
Глицеринфосфатдегидрогеназа
В печени – оба пути
Слайд 202). Образование ацил-КоА
путем активирования ВЖК (+ НSКоА)
путем новообразования из
ацетил-КоА (см. выше)
Слайд 213). Синтез фосфатидной кислоты
Глицерофосфатацилтрансфераза
1-ацилглицерол-3-фосфатацилтрансфераза
Слайд 224). Гидролиз фосфатидной кислоты
Фосфатидатфосфогидролаза
Слайд 235). Этерификация диглицерида
Диглицеридацилтрансфераза
Слайд 243. Из диоксиацетонфосфата
ДОАФ-ацилтрансфераза
1-ацилгидроксиацетонфосфатредуктаза
Относительно значения этого пути нет
единого мнения. По-видимому, он играет важную роль в пероксисомах, где
участвует в биосинтезе липидов с простой эфирной связью
Слайд 26Синтез наиболее важных фосфатидов локализован главным образом в ЭПС клетки
Слайд 271. Биосинтез лецитина
Образование ЦДФ-холина
1) холинкиназа
2) ЦТФ:холинфосфатцитидилилтрансфераза
Слайд 28Образование лецитина:
диглицерид + ЦДФ-холин лецитин + ЦМФ
Холинфосфотрансфераза
Самостоятельно: синтез кефалина
(коламинкиназа и коламинфосфотрансфераза)
1. Биосинтез лецитина
Слайд 29Образование ЦДФ-диглицерида
2. Синтез других фосфатидов
Фосфатидатцитидилилтрансфераза
Слайд 30Взаимодействие ЦДФ-диглицерида с азотистым основанием (сер или инозитом)
2. Синтез
других фосфатидов
Фосфатидилсеринсинтаза
Слайд 31Декарбоксилирование серинфосфатида
2. Синтез других фосфатидов
Фосфатидилсериндекарбоксилаза
Слайд 32Метилирование кефалина
кефалин + 3 S-аденозилметионин
лецитин + 3
S-аденозилгомоцистеин
2. Синтез других фосфатидов
Слайд 34Биосинтез холестерина
Примерно ½ холестерина, имеющегося в организме, образуется путем биосинтеза
в печени (50%), кишечнике (15%), коже – около 500 мг/сут.
Все клетки,
не утратившие ядро, способны синтезировать холестерин
Биосинтез его происходит в ЭПС и цитозоле клеток
Источник всех атомов С – ацетил-КоА
Синтез идет в несколько стадий
Слайд 351. Синтез мевалоновой кислоты из ацетил-КоА
Слайд 362. Образование изопреноидной единицы – изопентенилпирофосфата
Слайд 384. Циклизация сквалена с образованием ланостерола
Слайд 395. Превращение ланостерола в холестерин



домен")

 СН3–СОSКоА + НS–АПД НSКоА + СН3–СОSАПД")
 СН3–СО–СН2 –СОSАПД + НАДФН2 СН3–СН(ОН)–СН2–СОSАПД +")
 СН3–(СН2)14–СОS–АПД + Н2О СН3–(СН2)14–СООН + НS–АПД пальмитил-АПДТиоэстераза")



. Образование -фосфоглицерина 2 пути:фосфорилирование глицерина (в почках и у микроорганизмов)Глицеринкиназа")
. Образование -фосфоглицерина 2 пути:восстановление ДОАФ (в мышцах, в слизистой кишечника")
. Образование ацил-КоА путем активирования ВЖК (+ НSКоА)путем новообразования из ацетил-КоА (см. выше)")
. Синтез фосфатидной кислоты Глицерофосфатацилтрансфераза 1-ацилглицерол-3-фосфатацилтрансфераза")

 холинкиназа2) ЦТФ:холинфосфатцитидилилтрансфераза")

 2. Синтез других фосфатидов Фосфатидилсеринсинтаза")




2) Карбоксибиотин + СН3СОSКоА → → НООС–СН2–СОSКоА + биотин Малонил-КоА")

 моль1 мольацетил-КоАn мольмалонил-КоАВЖКС2(n+1)")

 1. Из -моноглицеридов")
. Гидролиз фосфатидной кислоты Фосфатидатфосфогидролаза")
. Этерификация диглицерида Диглицеридацилтрансфераза")