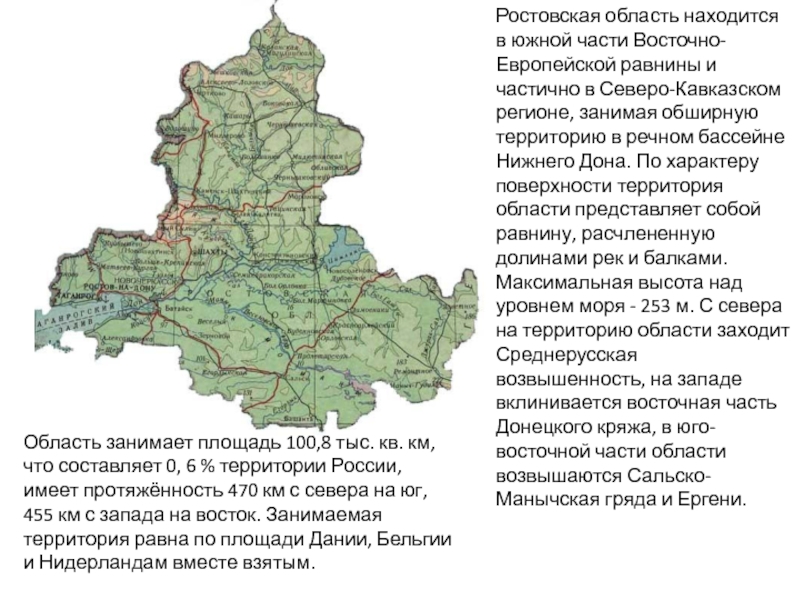
пуриновых оснований
Биосинтез пиримидиновых нуклеотидов
- синтез de novo
- регуляция биосинтеза
- реутилизация
пиримидиновых основанийРаспад пуриновых и пиримидиновых фнуклеотидов
Нарушения метаболизма нуклеотидов

Синтез нуклеотидов de novoАктивированная рибозаНуклеотидАктивированная рибозааминокислотыосно-ваниеНуклеотид")
 - ИНОЗИНМОНОФОСФАТ+ HCO3- UMP (УМФ) - УРИДИНМОНОФОСФАТ")
 Robert Greenberg (University")

+ATP+AMPРибозофосфатпирофосфокиназа (ФРПФ-синтетаза)Рибозо-5-фосфат5-фосфо-α-D-рибозил-1-пирофосфат (ФРПФ, PRPP)")

АИР-карбоксилазаКАИР-синтетазаКАИР-мутазаCAIR")




: аденин + PRPP → AMP + PPiГипоксантин-гуанинрибозилтрансфераза (КФ 2.4.2.8): гипоксантин гуанин")




уридин 5’-фосфат (уридилат, UMP)оротат-фосфорибозилтрансфераза оротидилатдекарбоксилаза")
")













В обеих реакциях окисляется NADPH.Высшие организмы должны получать фолиевую кислоту (фолат) с пищей")

НуклеозидыАзотистые основания:Урацил, тиминГуанин, гипоксантин (из аденина)Рибозо-1-фосфатДезоксирибозо-1-фосфат")





Избыточное образование мочевой кислоты↓Повышенное содержание мочевой кислоты в сыворотке↓Отложение кристаллов")

Почти полное отсутствие гипоксантин-гуанин-фосфорибозилтрансферазы ↓Повышенная концентрация ФРПФУвеличенная")