Разделы презентаций
- Разное
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Геометрия
- Детские презентации
- Информатика
- История
- Литература
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Обмен углеводов
Содержание
- 1. Обмен углеводов
- 2. Катаболизм основных пищевых веществ1-3 пищеварение; 4-8 специфические пути катаболизма;9-10 заключительный (общий путь) катаболизма;11- ЦПЭ;12- окислительное фосфорилирование
- 3. Аденозинтрифосфорная кислота (АТФ)
- 4. Обмен углеводов
- 5. Обмен углеводовПлан лекции:Функции углеводов в организмеОсновные углеводы
- 6. Строение и функции углеводов
- 7. Функции: - основные поставщики энергии (50% калорий
- 8. Основные углеводы пищиМоносахариды (Разобрать аномеры глюкозы) ОлигосахаридыПолисахариды… и
- 9. Превращение углеводов в желудочно-кишечном тракте В ЖКТ
- 10. Основное переваривание углеводов происходит в тонком кишечнике, где
- 11. Пищевые волокна (неперевариваемые углеводы): целлюлоза, пектиновые вещества,
- 12. Переваривание углеводов
- 13. Транспорт моносахаридов 1. Из просвета кишечника моносахариды
- 14. Рис. Всасывание моносахаридов в кишечнике
- 15. Облегченная диффузия – означает без затрат энергии.Следовательно,
- 16. Особенность ГЛЮТ 4, в том, что они
- 17. Метаболизм глюкозы в клеткахПосле поступления глюкозы в
- 18. Различают 2 фермента осуществляющие эту реакцию. В
- 19. Глюкокиназа (Км >10 мМ/л) печени (и мышц)
- 20. Ключевая роль глюкозо – 6-фосфата
- 21. В зависимости от физиологического состояния организма и
- 22. Схема катаболизма глюкозы
- 23. Катаболизм глюкозыГликолиз - это серия реакций, в
- 24. Таким образом, аэробный распад глюкозы это предельное
- 25. Гликолизсложный анаэробный ферментативный процесс, в ходе которого
- 26. Две стадии гликолизаI стадия гексоз В которой АТФ затрачиваетсяII стадия триоз образование АТФ
- 27. Последовательность реакций гликолиза
- 28. Реакции 1-й стадии гликолиза
- 29. Первая стадия гликолиза включает реакции превращения молекулы
- 30. В большинстве тканей реакцию фосфорилирования глюкозы катализирует
- 31. Продукт реакции изомеризации подвергается повторному фосфорилированию за
- 32. На следующем этапе фруктозо-1,6-дифосфат подвергается расщеплению на
- 33. Далее происходит изомеризация триозофосфатов (рис., реакция 5).
- 34. Реакции 2-й стадии гликолиза
- 35. Лактат по другому (по русски) молочная кислота
- 36. Вторая стадия гликолиза включает реакции превращения двух
- 37. 1,3-Дифосфоглицерат вступает в реакцию первого субстратного фосфорилирования,
- 38. Биологическое значение гликолизазаключается в образовании макроэргических соединений.
- 39. Суммарный эффект аэробного гликолиза составляет 8 моль
- 40. Анаэробный гликолизКонечный метаболит анаэробного гликолиза - лактат
- 41. Значение анаэробного гликолиза Анаэробный и аэробный гликолиз
- 42. Анаэробный гликолиз это брожениеРазличают несколько типов брожения,
- 43. Анаэробный гликолиз, несмотря на небольшой энергетический эффект,
- 44. Общие пути катаболизмаОкисление продуктов окисления глюкозы гликолизом
- 45. Картакатаболизма
- 46. Окислительное декарбоксилирование пировиноградной кислоты Суммарный результат многостадийной реакции выглядит следующим образом:
- 47. Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в пируватдегидрогеназный комплекс (ПДК):
- 48. Этот комплекс ферментов работает подобно конвейеру, в
- 49. Дигидролипоат-ацетилтрансфераза (2) - второй фермент комплекса. Катализирует
- 50. Дегидрогеназа дигидролипоевой кислоты (3й фермент) отщепляет водород
- 51. Цикл лимонной кислотыЦитратный цикл (цикл Кребса, цикл
- 52. В ходе одного полного цитратного цикла происходит:
- 53. Цикл лимонной кислоты / цикл Кребса
- 54. Энергетика полного окисления глюкозы в клетке1. Гликолиз
- 55. Ферменты цитратного цикла 1 – цитратсинтаза, 2 – аконитаза, 3
- 56. Сопряжение общих путей катаболизма с дыхательной цепьюВ
- 57. Энергетика цитратного цикла и общих путей катаболизма
- 58. Кроме того, в цитратном цикле происходит одна
- 59. Схема взаимосвязи общего пути катаболизма и ЦПЭ
- 60. задание на дом: записать все реакции ПДК и цикла Кребса Благодарю за внимание
- 61. Слайд 61
- 62. Пентозофосфатный путь в метаболизме глюкозы Это путь
- 63. Кроме того, пентозофосфатный путь (окислительный и неокислительный)
- 64. Пентозофосфатный путь превращений глюкозы
- 65. Пентозофосфатный путь является альтернативным путем окисления
- 66. 3глюкозо-6-фосфат + 6 НАДФ+ →→ 3СО2 +
- 67. Использование метаболитов ЦТК в синтезе различных соединений
- 68. Метаболизм глюкозы
- 69. Схема взаимосвязи общего пути катаболизма и ЦПЭ
- 70. Скачать презентанцию
Катаболизм основных пищевых веществ1-3 пищеварение; 4-8 специфические пути катаболизма;9-10 заключительный (общий путь) катаболизма;11- ЦПЭ;12- окислительное фосфорилирование
Слайды и текст этой презентации
Слайд 1…связь с обменом веществ, общие пути катаболизма поставщики субстратов для окислительного фосфорилирования Обмен
углеводов

Слайд 2Катаболизм основных пищевых веществ
1-3 пищеварение;
4-8 специфические пути катаболизма;
9-10 заключительный
(общий путь) катаболизма;
11- ЦПЭ;
12- окислительное фосфорилирование
 катаболизма;11- ЦПЭ;12- окислительное фосфорилирование")
")

Слайд 5Обмен углеводов
План лекции:
Функции углеводов в организме
Основные углеводы пищи
Пути поступления и
превращения углеводов в ткани организма
Транспортеры глюкозы: виды, особенности структуры и
функцииКлючевая роль глюкозо-6-фосфата, пути обмена.
Гликолиз – центральный путь катаболизма глюкозы и моносахаридов.
Лактат и пируват: пути обмена, значение, реакции превращения в Ацетил-КоА и ЩУК. Энергетический баланс окисления до СО2 и Н2О.


Слайд 7Функции:
- основные поставщики энергии (50% калорий в сутки),
-
резервная (гликоген)
- структурная (гликозаминогликаны межклеточного матрикса, компоненты соединительной ткани),
- входят в состав и участвуют в функциях гликопротеинов (ферментов, белков -транспортёров, белков-рецепторов, иммуноглобулинов, гормонов),
- являются компонентами мембран эритроцитов определяющих группу крови,
- входят в состав коферментов и нуклеиновых кислот,
- входят в состав гликолипидов мембран,
- необходимы для обезвреживания вредных веществ.
, - резервная")
Слайд 8Основные углеводы пищи
Моносахариды (Разобрать аномеры глюкозы)
Олигосахариды
Полисахариды
… и декстрины
Объяснить сокращения:
Гли, глю,
гал, фру, ман…
Нормы потребления углеводов в сутки (500 г).
 ОлигосахаридыПолисахариды… и декстриныОбъяснить сокращения: Гли, глю, гал,")
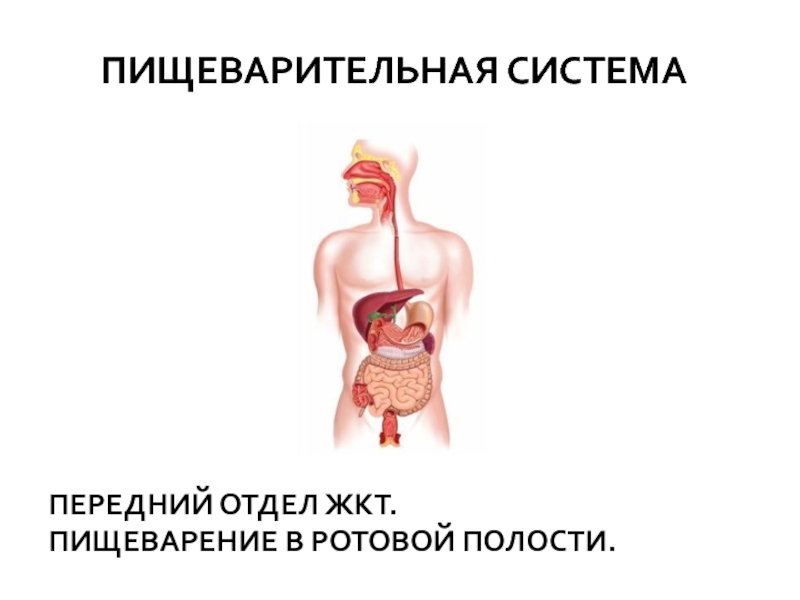
Слайд 9Превращение углеводов в
желудочно-кишечном тракте
В ЖКТ основной углеводный элемент пищи
крахмал начинает расщепляться в ротовой полости до декстринов
и мальтозы
под действием содержащейся в слюне α-амилазы - фермента класса гидролаз.
Т. к. время пребывания пищи в ротовой полости мало, процесс переваривания там незначителен.
В полости желудка распада углеводов не происходит,
так как в нём не вырабатываются соответствующие
ферменты. Оптимум действия α-Амилаза слюны
рН=6,8-7,2 и она неактивна в кислом содержимом желудка.

Слайд 10 Основное переваривание углеводов происходит в тонком
кишечнике, где имеются соответствующие ферменты
и
слабощелочная среда.
В тонком кишечнике работают амилазы поджелудочной железы
В
тонком кишечнике рН 6,7-7,2, что обеспечивает их высокую активность. α-Амилаза поджелудочной железы гидролизует крахмал
и гликоген до декстринов, а затем до мальтозы.
α–1,6-гликозидную связь в крахмале гидролизуют
ферменты амило-1,6-глюкозидаза и
олиго-1,6-глюкозидаза.
Гликозидные связи в дисахаридах гидролизуют
специфические гидролазы тонкого кищечника:

Слайд 11
Пищевые волокна (неперевариваемые углеводы): целлюлоза, пектиновые вещества, хитин в
ЖКТ
не расщепляются, но необходимы организму, так
как способствуют продвижению пищи
в кишечнике и выводят токсичные вещества и продукты распада из
организма.
Моносахариды через стенки кишечника поступают в
кровь. Глюкоза и галактоза всасываются значительно
быстрее, чем другие монозы.
: целлюлоза, пектиновые вещества, хитин в ЖКТ не")

Слайд 13Транспорт моносахаридов
1. Из просвета кишечника моносахариды в клетки слизистой
кишечника (энтероциты) осуществляется путём:
- облегчённой диффузии и
- активного транспорта.
Для
транспорта глюкозы необходимы ионы Na+2. Из клеток кишечника глюкоза перемещается во внеклеточную жидкость и далее в кровь с помощью облегченной диффузии.
3. Далее по воротной вене глюкоза попадает в печень, где часть её задерживается, а другая (меньшая) часть поступает в клетки других органов и тканей путем облегченной диффузии при участии белков-переносчиков транспортеров глюкозы (ГЛЮТ).


Слайд 15Облегченная диффузия – означает без затрат энергии.
Следовательно, скорость трансмембранного потока
глюкозы из крови в клетки тканей и органов зависит
только
от градиента её концентрации. Исключением являются клетки мышц и жировой ткани,
где облегченная диффузия регулируется гормонами
инсулином и глюкагоном.
Разновидности ГЛЮТ:
ГЛЮТ 1 – в мозге,
ГЛЮТ 2 – в печени и поджелудочной железе,
ГЛЮТ 3 – во многих тканях, и в печени и в почках,
ГЛЮТ 4 – в мышцах и в жировой ткани,
ГЛЮТ 5 – в тонкой кишке

Слайд 16Особенность ГЛЮТ 4, в том, что они находятся в
цитоплазме
клеток мышц и адипоцитов в визикулах
(пузырьках). Влияние инсулина на такие
клетки приводит к перемещению визикул, содержащих ГЛЮТ 4
к плазматической мембране и их слиянию с ней, после
чего возможен облегченный транспорт глюкозы в эти
клетки. После снижения концентрации инсулина в крови
белки-транспортеры глюкозы снова перемещаются в
цитозоль.
Печень инсулин независимая ткань.
Почки в здоровом организме глюкозу из первичной мочи
полностью реабсорбируют

Слайд 17Метаболизм глюкозы в клетках
После поступления глюкозы в клетки всех тканей
происходит
её фосфорилирование:
Глюкоза + АТФ = Глюкозо-6-фосфат + АДФ
Плазматическая мембрана клеток
непроницаема для Глю-6-ф и глюкоза из клетки уже выйти не может.

Слайд 18Различают 2 фермента осуществляющие эту реакцию.
В печени и поджелудочной
железе Глюкокиназа
Во всех других тканях Гексокиназа
Эти ферменты отличаются константой
Км.Км Гюкокиназы (>10 мМ/л) → низкое сродство
Км Гексокиназы (< 0,1 мМ/л) → высокое сродство
Вследствие этого:
мозг, эритроциты и др. ткани (с ферментом Гексокиназой
(Км < 0,1 мМ/л) могут использовать глюкозу при
снижении её концентрации в крови в постсорбтивный
период (через 4-5 часов после еды) и при голодании.

Слайд 19Глюкокиназа (Км >10 мМ/л) печени (и мышц)
активно использует глюкозу
только при её высокой
концентрации в крови в абсорбтивный период
(приалиментарной гипергликемии) и ограничивает - при
снижении её уровня в крови.
Алиментарная гипергликемия – повышение
концентрации глюкозы в крови после приёма пищи
Через 0,5 – 1 часа после приема пищи и до 2 часов.
 печени (и мышц) активно использует глюкозу только")

Слайд 21В зависимости от физиологического состояния организма и типа
ткани глюкозо-6-фосфат
может использоваться в разных
метаболических процессах.
1 – в большинстве тканей
синтезируется гомополисахарид гликоген2 – во многих тканях используется для синтеза аминокислот,
3 – в адипоцитах и гепатоцитах используется на построение жиров,
4 – в эритроцитах и скелетных мышцах может превращается в лактат.
5 – в разных тканях используется для построения других
моносахаридов и обязательно используется как источник
энергии


Слайд 23Катаболизм глюкозы
Гликолиз - это серия реакций, в результате
которых глюкоза
распадается на две молекулы
пирувата (аэробный гликолиз) или две молекулы
лактата (анаэробный гликолиз).
Все 10 реакций гликолиза протекают в
цитозоле клеток (цитоплазме) и характерны для
всех органов и тканей.
Аэробный распад глюкозы включает реакции
аэробного гликолиза и последующее окисление
пирувата в реакциях катаболизма.

Слайд 24Таким образом, аэробный распад глюкозы
это предельное ее окисление до
СО2 и Н2О,
а анаэробный гликолиз - это
специфический путь
катаболизма, то есть часть аэробного распада глюкозы.
Анаэробный распад включает те же реакции
специфического пути распада глюкозы до
пирувата, но с последующим превращением
пирувата в лактат
(то есть термины анаэробный распад и
анаэробный гликолиз совпадают).
Последовательность реакций гликолиза
приведена на рисунке:

Слайд 25Гликолиз
сложный анаэробный ферментативный процесс, в ходе
которого молекула глюкозы превращается
в 10-ти
последовательных реакциях в 2 молекулы пировиноградной
кислоты (ПВК).
Процесс гликолиза катализируется 10 ферментами и
протекает в цитоплазме клеток.
В этом процессе часть энергии запасается в виде АТФ.
Процесс гликолиза универсален: окисление глюкозы
протекает одинаково у растений, животных,
микроорганизмов и т.д. и различается только
характером регуляции скорости процесса и судьбой
образованной ПВК.
Условно процесс можно разделить на 2 стадии.

Слайд 26Две стадии гликолиза
I стадия гексоз
В которой АТФ затрачивается
II стадия триоз
образование
АТФ



Слайд 29Первая стадия гликолиза включает реакции превращения молекулы глюкозы в две
молекулы фосфотриоз. Эта стадия сопровождается затратой молекул АТФ.
Начальной реакцией
глюкозы в клетке является её фосфорилирование в результате взаимодействия с АТФ (рис., реакция 1). Эта реакция в условиях клетки протекает только в одном направлении. Биологическая роль реакции фосфорилирования глюкозы заключается в том, что глюкозо-6-фосфат, в отличие от свободной глюкозы, не может проникать через клеточную мембрану обратно в кровь. 
Слайд 30В большинстве тканей реакцию фосфорилирования
глюкозы катализирует фермент гексокиназа, которая
обладает высоким сродством к глюкозе, способна также
фосфорилировать фруктозу и маннозу
и ингибируетсяизбытком глюкозо-6-фосфата.
В клетках печени, есть фермент глюкокиназа, которая имеет низкое сродство к глюкозе, не ингибируется глюкозо-6-фосфатом и не участвует в фосфорилировании других моносахаридов.
В следующей реакции глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат (рис., реакция 2).

Слайд 31Продукт реакции изомеризации
подвергается повторному
фосфорилированию за счёт АТФ
(рис.
реакция 3).
Эта реакция – наиболее медленно протекающая реакция
гликолиза и,
подобно фосфорилированию глюкозы, необратима.
Фермент – фосфофруктокиназа – является
аллостерическим, активируется АДФ и
АМФ, ингибируется цитратом и высокой
концентрацией АТФ.

Слайд 32На следующем этапе фруктозо-1,6-дифосфат
подвергается расщеплению на две фосфотриозы
(рис.
реакция 4).
Таким образом, химическое соединение,
содержащее 6 углеродных атомов, превращается
в два, содержащих по 3 атома углерода.
Поэтому гликолиз называют дихотомическим путём
превращения глюкозы
(от слова дихотомия – рассечение на две части).

Слайд 33Далее происходит изомеризация триозофосфатов
(рис., реакция 5).
В этой реакции
диоксиацетонфосфат переходит в
глицеральдегид-3-фосфат.
Таким образом, в первой стадии гликолиза
молекула глюкозы
превращается в две молекулы глицеральдегид-3-фосфата.
. В этой реакции диоксиацетонфосфат")

 молочная кислота")
Слайд 36Вторая стадия гликолиза включает реакции превращения двух молекул глицеральдегид-3-фосфата в
две молекулы пировиноградной кислоты (ПВ
А в анаэробном гликолизе лактата.
На
этой стадии гликолиза происходит синтезмолекул АТФ.
Глицеральдегид-3-фосфат подвергается дегидрированию при участии НАД-зависимой дегидрогеназы. В этой реакции происходит потребление неорганического фосфата, который включается в состав продукта реакции, содержащего макроэргическую фосфатную связь (рис., реакция 6).

Слайд 371,3-Дифосфоглицерат вступает в реакцию
первого субстратного фосфорилирования,
т.е. не
сопряжённого с переносом электронов в
дыхательной цепи митохондрий.
В
этой реакции осуществляется синтез молекулы АТФ в результате переноса фосфатной группы вместе с
макроэргической связью на молекулу АДФ
(рис., реакция 7).
Реакции 8-10 объяснить. 11- = лактат.

Слайд 38Биологическое значение гликолиза
заключается в образовании макроэргических соединений.
В первой стадии
гликолиза затрачиваются две молекулы
АТФ (реакции 1 и 3).
Во
второй стадии гликолиза из двух молекул триоз образуются четыре молекулы АТФ (реакции 7 и 10),
таким образом при превращении одной молекулы глюкозы
Образуются : 2 молекулы АТФ,
2 молекулы НАДН и
2 молекулы ПВК.

Слайд 39Суммарный эффект аэробного гликолиза
составляет 8 моль АТР, так как
в реакциях 1 и 3
используется 2 моль АТР.

Слайд 40Анаэробный гликолиз
Конечный метаболит анаэробного гликолиза - лактат
или молочная кислота
Все
анаэробные процессы энергетически более бедные чем аэробные.
В анаэробных процессах образуется
меньше АТФ
Слайд 41Значение анаэробного гликолиза
Анаэробный и аэробный гликолиз
энергетически неравноценны.
Образование
двух моль лактата из глюкозы
сопровождается синтезом всего двух моль
АТР, потому что НАДH+Н, полученный при окислении
глицероальдегидфосфата, не используется дыхательной
цепью, а акцептируется пируватом для синтеза
молочной кислоты (лактата) (11 реакция).

Слайд 42Анаэробный гликолиз это брожение
Различают несколько типов брожения, например:
молочнокислое брожение,
спиртовое
брожение
т.е. после образования в гликолизе пирувата,
пируват не поступает в
митохондрии и можетпревращаться или в молочную кислоту.
В каких клетках организма нет митохондрий?
Это конечный продукт окисления глюкозы многих
анаэробных микроорганизмов.

Слайд 43Анаэробный гликолиз,
несмотря на небольшой
энергетический эффект,
является основным
источником
энергии
для скелетных мышц в
начальном периоде
интенсивной работы, то
есть в условиях, когда
снабжение кислородом
ограничено. Кроме того,
зрелые эритроциты
извлекают энергию за счет
анаэробного окисления
глюкозы, потому что не
имеют митохондрий.

Слайд 44Общие пути катаболизма
Окисление продуктов окисления глюкозы гликолизом
ПВК в общих
путях катаболизма в митохондриях:
Пируват декарбоксилазным комплексом (ПДК) декарбоксилирование и дегидрирование
с образованием АцетилКоА.В цитратном цикле окисление АцКоА до СО2 и Н2О


Слайд 46Окислительное декарбоксилирование пировиноградной кислоты Суммарный результат многостадийной реакции выглядит следующим образом:

Слайд 47Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными
в пируватдегидрогеназный комплекс (ПДК):
:")
Слайд 48Этот комплекс ферментов работает подобно
конвейеру, в котором продукт передается
от
фермента к ферменту. Такой принцип повышает
эффективность работы ферментов, так
как снижаетслучайность в контакте реагирующих веществ с
ферментом. М.м. ПДК более 6х1000 000 Да
Далее приводятся названия ферментов и
характеристика катализируемых реакций.
Пируватдекарбоксилаза (1). В качестве кофермента в
реакции участвует тиаминдифосфат - производное
витамина В1.
Фермент катализирует отщепление карбоксильной группы
в виде СО2, а ацетильный остаток присоединяет к
липоевой кислоте - коферменту второго фермента.
Получается ацетил-липоат.

Слайд 49Дигидролипоат-ацетилтрансфераза (2) - второй фермент
комплекса. Катализирует перенос ацетильного остатка,
соединенного с липоевой кислотой на второй кофермент
HS-КоА
с образованием ацетил-КоА. Таким образом, в этой реакции участвуют два кофермента:
липоевая кислота, прочно соединенная с ферментом, и
кофермент А, объединяющийся с ферментом в момент
реакции. Водород остается связанным с липоевой кислотой,
которая превращается в дигидролипоат.
 - второй фермент комплекса. Катализирует перенос ацетильного остатка, соединенного")
Слайд 50Дегидрогеназа дигидролипоевой кислоты (3й фермент)
отщепляет водород от липоевой кислоты
и
переносит его на НАД+. Далее водород на НАДН
транспортируется в
дыхательную цепью.Главные продукты реакции - это
НАДH и ацетил-СоА.
NADH далее окисляется в дыхательной цепи,
где энергия используется на синтез 3 моль АТР, а
ацетил-СоА окисляется в цитратном цикле.
Пируватдекарбоксилазный комплекс находится на
внутренней мембране митохондрий и соединен с
ней со стороны матрикса.
 отщепляет водород от липоевой кислоты и")
Слайд 51Цикл лимонной кислоты
Цитратный цикл
(цикл Кребса, цикл трикарбоновых кислот (ЦТК)
–
это система реакций, приводящая к полному
окислению двухуглеродного ацетильного
фрагмента
до СО2 И Н2О, имеющего различное происхождение.
Цитратный цикл является общим конечным путем
окисления белков, жиров и углеводов.
Все реакции цитратного цикла,
как и окислительного декарбоксилирования
пирувата, локализованы в митохондриях.
 –это")
Слайд 52В ходе одного полного цитратного цикла происходит:
1. полное окисление
ацетильного остатка до двух молекул СО2;
2. образование трех молекул
восстановленного НАДН и одной молекулы ФAДH2; 3. образование одной молекулы ГТФ в результате субстратного фосфорилирования.
Реакции цитратного цикла,
ферменты и их характеристика приведена на
следующем слайде:


Слайд 54Энергетика полного окисления глюкозы в клетке
1. Гликолиз (распад глю до
2х ПВК) – 8 АТФ
2.
Пируват-декарбоксилазный комплекс 2 молекулы ПВК до 2 ацКоа 6 АТФ
3. 2 молекулы АцКоа до СО2 и Н2О 24 АТФ
----------------
Итого: 38 АТФ

Слайд 55Ферменты цитратного цикла
1 – цитратсинтаза,
2 – аконитаза,
3 – изоцитратдегидрогеназа,
4
– a-кетоглутаратдегидрогеназный комплекс,
5 – сукцинил-КоА-тиокиназа,
6 – сукцинатдегидрогеназа,
7 –
фумараза, 8 – малатдегидрогеназа.

Слайд 56Сопряжение общих путей катаболизма с дыхательной цепью
В общих путях катаболизма
происходит
пять реакций дегидрирования:
(одна на стадии окислительного декарбоксилирования
пирувата
и четыре в цитратном цикле). Все 10 атомов водорода переносятся на коферменты
дегидрогеназ, которые в свою очередь окисляются
в дыхательной цепи. Окисленные коферменты
возвращаются в реакции общих путей катаболизма.
Регенерация коферментов - это обязательное
условие для протекания реакции дегидрирования.
Таким образом, общий путь катаболизма
и дыхательная цепь непрерывно связаны между собой и
отдельно функционировать не могут.

Слайд 57Энергетика цитратного цикла и общих путей катаболизма
За один оборот
цитратного цикла синтезируется
12 молекул АТФ.
Девять из них образуются за
счет энергии транспорта в дыхательной цепи трех пар водорода от трех молекул НАДH + H+.Две молекулы АТР синтезируются при окислении 1 молекулы FADH2, так как в дыхательной цепи в данном случае действуют только два пункта сопряжения с окислительным фосфорилированием ADP.

Слайд 58Кроме того, в цитратном цикле происходит одна
реакция субстратного фосфорилирования,
дающая 1 моль ГТФ ( аналог АТР).
ИТОГО:
В общих путях катаболизма
синтезируется 15 молекул АТФ.
Три из них при окислительном декарбоксилировании пирувата (в ПДК) и
12 - в цитратном цикле.




Слайд 62Пентозофосфатный путь в метаболизме глюкозы
Это путь превращения глюкозы в
пентозы. В пентозофосфатном пути
превращения глюкозы можно выделить две
части:
А - окислительный путь и
Б - неокислительный путь синтеза пентоз.
Коферментом дегидрогеназ является НАДН+Н, который восстанавливается в НАДФH и используется клетками в реакции восстановления и гидроксилирования.

Слайд 63Кроме того,
пентозофосфатный путь (окислительный и
неокислительный)
поставляет клетке пентозофосфаты
(в основе рибоза и дезоксирибоза),
необходимые для синтеза нуклеиновых
кислот
и коферментов (НАД, НАДФ, ФМН и ФАД). Все реакции пентозофосфатного пути
проходят в цитозоле клетки. Реакции
неокислительного этапа пентозофосфатного
пути являются обратимыми, поэтому
становится возможным синтез гексоз из пентоз.
 поставляет клетке пентозофосфаты (в")

Слайд 65
Пентозофосфатный путь является
альтернативным путем окисления глюкозы.
Он включает
несколько циклов, в результате
функционирования которых из трех молекул
глюкозо-6-фосфата
образуются три молекулы СО2 и три молекулы пентоз.
Последние используются для регенерации двух
молекул глюкозо-6-фосфата и одной молекулы
глицеральдегид-3-фосфата. Поскольку из двух
молекул глицеральдегид-3-фосфата можно
регенерировать молекулу глюкозо-6-фосфата,
глюкоза может быть полностью окислена при
превращении по пентозофосфатному пути:

Слайд 663глюкозо-6-фосфат + 6 НАДФ+ →
→ 3СО2 + 2глюкозо-6-фосфат +
глицеральдегид-3-фосфат
+ 6 НАДФ-Н2
Пентозофосфатный цикл не приводит к синтезу
АТФ. Он выполняет
две главные функции:образование НАДФ-Н2 для восстановительных синтезов, таких, как синтез жирных кислот и синтез стероидов (в цитоплазме клетки).
2) обеспечение рибозой синтеза нуклеотидов, синтеза нуклеиновых кислот и коферментов.