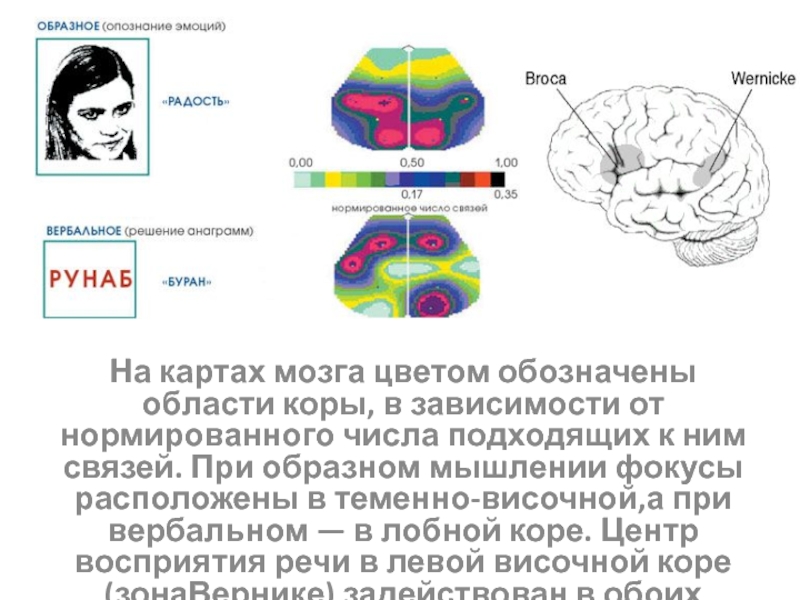
глікогену
Глікоген – великий розгалужений полімер, що складається із залишків глюкози.
Розщеплюється до глюкози, коли є потреба в енергіїСИНТЕЗ І РОЗЩЕПЛЕННЯ ГЛІКОГЕНУ

 і скелетні м’язи (2 %) – два")


Дві форми ГФ Фосфорилаза")




 в реакції, що каталізується глюкозо-1-фосфат уридилтрансферазою")



Підвищує швидкість")























Глюкоза, утворена")