Разделы презентаций
- Разное
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Геометрия
- Детские презентации
- Информатика
- История
- Литература
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
MODULUL 9
Содержание
- 1. MODULUL 9
- 2. 9.1. Основы генной инженерииМатериально-техническая база генной инженерииОсновные этапы генной инженерии
- 3. Проверим наши знания: 1. ЧТО ЯВЛЯЕТСЯ ОСНОВНОЙ
- 4. 2. Сколько типов тРНК могут быть в
- 5. 3. Генетическая рекомбинация у бактерий осуществляется следующими
- 6. 4. Сколько хромосом отошло к полюсам клетки
- 7. 5. Наследование групп крови у человека в
- 8. 6. КАКИМИ СВЯЗЯМИ СВЯЗАНЫ МЕЖДУ СОБОЙ ЦЕПИ
- 9. 7. ПЛАЗМИДЫ ЯВЛЯЮТСЯ: A) молекулами ДНК
- 10. Проверим наши знания: 1. ЧТО ЯВЛЯЕТСЯ ОСНОВНОЙ
- 11. 2. Сколько типов тРНК могут быть в
- 12. 3. Генетическая рекомбинация у бактерий осуществляется следующими
- 13. 4. Сколько хромосом отошло к полюсам клетки
- 14. 5. Наследование групп крови у человека в
- 15. 6. КАКИМИ СВЯЗЯМИ СВЯЗАНЫ МЕЖДУ СОБОЙ ЦЕПИ
- 16. 7. ПЛАЗМИДЫ ЯВЛЯЮТСЯ: A) молекулами ДНК
- 17. Проверим полученные результаты !
- 18. Вернемся к нашим …….
- 19. Вернемся к нашим …….
- 20. Генетическая инженерияМанипуляция на уровне генома организмов с
- 21. Когда возникла генная инженерия?1972, P.BergПолучение первой рекомбинантной молекулы ДНК1944, A.Avery, C.McLeod, M.McCartyОткрытие явления трансформации
- 22. Основные этапы развития генной инженерии:Открытие основных механизмов
- 23. Основные этапы развития генной инженерии (продолжение):Открытие обратной
- 24. 2. Материально-техническая база генной инженерииГенная инженерия основывается
- 25. Ферменты (основные группы рестриктаз)Endonucleaze Pentru activitate necesită
- 26. Свойства рестриктазМогут разрезать молекулу ДНК на дискретные
- 27. “Site”-узнавания некоторых рестриктаз:
- 28. Примеры рестрикции Recognition Site
- 29. Название ферментов рестрикцииНазвание рестриктаз происходит от названия
- 30. Векторные системыСпециализированные молекулы по переносу наследственной информации
- 31. Требования предъявляемые векторным системамБезвредность вектораБыстрое размножение в
- 32. Использование векторовПлазмиды Для клонирования ДНК которые содержат
- 33. Клетки хозяева (требования)Возможность вовлечения в исследовательский процессОтсутствие
- 34. 3. Этапы генной инженерии1. Изолирование и /
- 35. 1. Получение генов1.1. Изолирование геновГибридизация ДНК –
- 36. 1. Получение генов1.2. Синтез геновИз смеси дезоксиполинуклеотидтрифосфатов
- 37. 2. Получение рекомбинарнтных молекул ДНК1.1. лигазный методИспользование
- 38. 3. Перенос и экспрессия рекомбинантных молекул ДНКТипы
- 39. Механизмы защиты экзогенной генетической информацииМетилирование ДНК (вирусы)Перевод
- 40. Механизмы защиты эндогенной генетической информацииМетилирование ДНКДействие специфических
- 41. ..... ?! .....КТО КОГО?!
- 42. Скачать презентанцию
9.1. Основы генной инженерииМатериально-техническая база генной инженерииОсновные этапы генной инженерии
Слайды и текст этой презентации

Слайд 29.1. Основы генной инженерии
Материально-техническая база генной инженерии
Основные этапы генной инженерии

Слайд 3Проверим наши знания:
1. ЧТО ЯВЛЯЕТСЯ ОСНОВНОЙ ЕДИНИЦЕЙ НАСЛЕДСТВЕННОСТИ?
A) аминокислота
белок аминокислота")
 4")
Слайд 5
3. Генетическая рекомбинация у бактерий осуществляется следующими способами:
1.
репликация
2. трансформация
3. трансляция
4. трансдукция
5. конъюгацияA) 1, 2, 4, 5 B) 2, 3, 4
C) 2, 4, 5 D) только 2 и 4

Слайд 6
4. Сколько хромосом отошло к полюсам клетки в анафазе I
мейоза, если исходная клетка содержала 14 хромосом?
A) 0
B) 7
C) 14
D) 28

Слайд 7
5. Наследование групп крови у человека в система АВО:
1. определяется взаимодействием аллельных генов типа множественного аллелизма
2.
определяется взаимодействием аллельных генов типа кодоминирования3. определяется взаимодействием аллельных генов типа плейотропии
4. определяется взаимодействием неаллельных генов типа комплемантарии
5. определяется взаимодействием аллельных генов типа эпистазии
A) 1, 3 B) 1, 5
C) 1, 2, 5 D) 1, 3, 5 E) 2, 5

Слайд 8
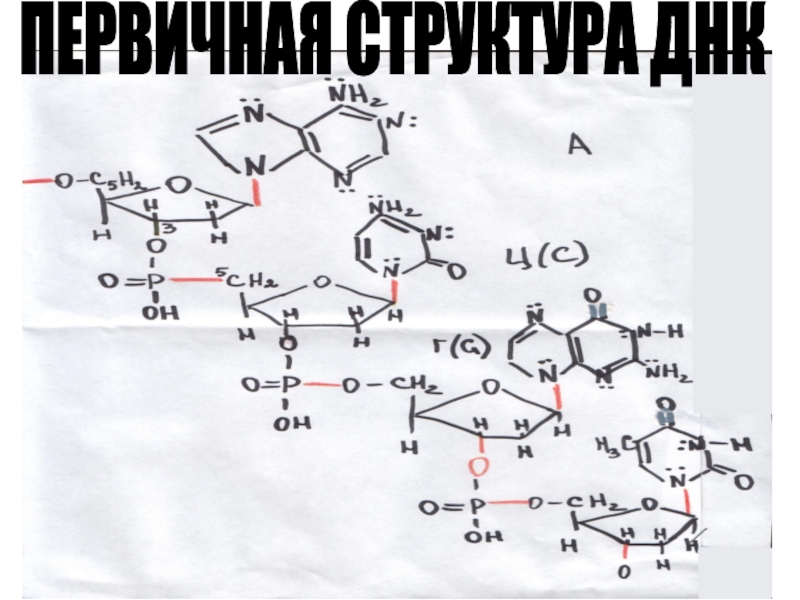
6. КАКИМИ СВЯЗЯМИ СВЯЗАНЫ МЕЖДУ СОБОЙ ЦЕПИ В МОЛЕКУЛЕ ДНК?
A) пептидными
B) водородными
C) фосфодиэфирными
D) сульфиднымиE) все варианты являются правильными
")
Слайд 9
7. ПЛАЗМИДЫ ЯВЛЯЮТСЯ:
A) молекулами ДНК с экзон-интронной
организацией
B) бактериальными молекулами
ДНК, которые не могут передаваться
другим клеткам
C) кольцевыми молекулами ДНК
D) молекулами ДНК, которые
реплицируются под непосредственным
контролем бактериальной хромосомы
 молекулами ДНК с экзон-интронной")
Слайд 10Проверим наши знания:
1. ЧТО ЯВЛЯЕТСЯ ОСНОВНОЙ ЕДИНИЦЕЙ НАСЛЕДСТВЕННОСТИ?
A) аминокислота
B) нуклеотид
C) ген
D)
белок аминокислота")
 4")
Слайд 12
3. Генетическая рекомбинация у бактерий осуществляется следующими способами:
1.
репликация
2. трансформация
3. трансляция
4. трансдукция
5. конъюгацияA) 1, 2, 4, 5 B) 2, 3, 4
C) 2, 4, 5 D) только 2 и 4

Слайд 13
4. Сколько хромосом отошло к полюсам клетки в анафазе I
мейоза, если исходная клетка содержала 14 хромосом?
A) 0
B) 7
C) 14
D) 28

Слайд 14
5. Наследование групп крови у человека в система АВО:
1. определяется взаимодействием аллельных генов типа множественного аллелизма
2.
определяется взаимодействием аллельных генов типа кодоминирования3. определяется взаимодействием аллельных генов типа плейотропии
4. определяется взаимодействием неаллельных генов типа комплемантарии
5. определяется взаимодействием аллельных генов типа эпистазии
A) 1, 3 B) 1, 5
C) 1, 2, 5 D) 1, 3, 5 E) 2, 5

Слайд 15
6. КАКИМИ СВЯЗЯМИ СВЯЗАНЫ МЕЖДУ СОБОЙ ЦЕПИ В МОЛЕКУЛЕ ДНК?
A) пептидными
B) водородными
C) фосфодиэфирными
D) сульфиднымиE) все варианты являются правильными
")
Слайд 16
7. ПЛАЗМИДЫ ЯВЛЯЮТСЯ:
A) молекулами ДНК с экзон-интронной
организацией
B) бактериальными молекулами
ДНК, которые не могут передаваться
другим клеткам
C) кольцевыми молекулами ДНК
D) молекулами ДНК, которые
реплицируются под непосредственным
контролем бактериальной хромосомы
 молекулами ДНК с экзон-интронной")



Слайд 20Генетическая инженерия
Манипуляция на уровне генома организмов с целью их совершенствования
Пр.:
экспериментальный мутагенез, гибридизация
Генная инженерия
- Манипуляция на уровне ДНК с целью
создания рекомбинантных молекул и их переноса в другие организмы- Пр.: получение гормонов, получение генетически модифицированных организмов (ГМО)

Слайд 21Когда возникла генная инженерия?
1972, P.Berg
Получение первой рекомбинантной молекулы ДНК
1944, A.Avery,
C.McLeod, M.McCarty
Открытие явления трансформации

Слайд 22Основные этапы развития генной инженерии:
Открытие основных механизмов переноса генетической информации
Трансформация
(Griffits, 1928; Avery, McLeod, McCarty, 1944)
Сексдукция (Lederberg, Tatum, 1946)
Трансдукция (Tsinder,
Lederberg, 1952)Открытие структуры ДНК (Watson, Crick, 1953)
Открытие основных ферментов (50-60 гг.)
Рестриктазы
Лигазы

Слайд 23Основные этапы развития генной инженерии (продолжение):
Открытие обратной транскрипции (G.Temin, S.Mizutani,
D.Baltimor, 1970)
Открытие возможности искусственного синтеза генов (H.Corana, 1970-76)
Получение первой рекомбинантной
молекулы ДНК (P.Berg, 1972)Искусственное получение инсулина (1982)
:Открытие обратной транскрипции (G.Temin, S.Mizutani, D.Baltimor,")
Слайд 242. Материально-техническая база генной инженерии
Генная инженерия основывается на изолирование отдельных
фрагментов ДНК и их клонирование в векторные системы которые реплицируются
автономно в клетки (организмы) хозяеваОна включает:
Систему ферментов
Систему переноса (векторы)
Клетки хозяева

Слайд 25Ферменты (основные группы рестриктаз)
Endonucleaze
Pentru activitate necesită prezenţa în mediu
de incubare ATP, S-adenozilmetionină, Mg2+
Sunt instabile
Nu produc fragmente discrete
“site”- специфические эндонуклеазы
Для активности нуждаются лишь в Mg2+
Около 500
Производят дискретные фрагменты, легко выделяются электрофорезом в агарозном или полиакриламидном геле
Grupa intermediară
Pentru activitate necesită prezenţa a Mg2+ şi ATP
Produc fragmente discrete
Endonucleaze Pentru activitate necesită prezenţa în mediu de")
Слайд 26Свойства рестриктаз
Могут разрезать молекулу ДНК на дискретные фрагменты
Относительно легко выделяются
Способность образовывать “липкие концы” (не все рестриктазы)
Высокая специфичность действия (разрезает
молекулу ДНК в определенных местах – сайтах узнавания)

Слайд 28Примеры рестрикции
Recognition Site Cleavage
Products
EcoR1 ---GAATTC---
---G AATTC-----CTTAAG--- ---CTTAA G—
STICKY ENDS
Sma 1 ---CCCGGG--- ---CCC GGG---
---GGGCCC--- ---GGG CCC---
BLUNT ENDS
Участки рестрикции содержат 4 – 8 пар оснований в длину

Слайд 29Название ферментов рестрикции
Название рестриктаз происходит от названия бактерий из которых
они выделены:
Пр.: EcoRI
Из E.coli, штамм R
I – число (первый)
выделенного фермента
Слайд 30Векторные системы
Специализированные молекулы по переносу наследственной информации рекомбинантных молекул ДНК
в клетки (организмы) хозяева
Примеры:
- плазмиды
- фаги
- вирусы
- космиды
- митохондриальное
ДНК / хлоропластное ДНК
Слайд 31Требования предъявляемые векторным системам
Безвредность вектора
Быстрое размножение в клетке-хозяине и разрушение
вне ее
Наличие ограниченного количества сайтов рестрикции
Наличие маркерных генов для узнавания
Относительная
легкость выделенияКлонирование и экспрессия в другие клетки

Слайд 32Использование векторов
Плазмиды
Для клонирования ДНК которые содержат от нескольких пар
оснований (pb) до нескольких тысяч
Фаги
Для клонирования ДНК которые содержат
10.000 – 20.000 пар оснований (pb)Искусственные “хромосомы”
Для клонирования ДНК которые содержат сотни тысяч пар оснований (pb)

Слайд 33Клетки хозяева (требования)
Возможность вовлечения в исследовательский процесс
Отсутствие возможности существовать вне
лабораторных условиях
Возможность разрушения ДНК вне лабораторных условиях
Исключение возможности передачи
наследственной информации другим организмамИсключение возможности биологического загрязнения окружающей среды
Возможность вовлечения в исследовательский процессОтсутствие возможности существовать вне лабораторных")
Слайд 343. Этапы генной инженерии
1. Изолирование и / или синтез генов
2.
Клонирование генов и получение рекомбинантных молекул ДНК
3. Перенос и экспрессия
рекомбинантных молекул ДНК в клетки (организмы) хозяева
Слайд 351. Получение генов
1.1. Изолирование генов
Гибридизация ДНК – ДНК (Dj.Shapiro et
al., 1969)
гибридизация фагов λ и φ80;
изолирование участка lac-оперона;
Гибридизация ДНК –
РНК (Tomas, 1976)- комплекс ДНК – РНК является более устойчивым чем ДНК – ДНК;
Гибридизация РНК – ДНК (G.Temin, B.Mizutani, D.Baltimor, 1970)
- использование реверстранскриптаз;
Фрагментация ДНК (Maniatis et al., 1978)
используется для ДНК эукариот;
предполагает анализ фрагментов на селективных средах

Слайд 361. Получение генов
1.2. Синтез генов
Из смеси дезоксиполинуклеотидтрифосфатов в присутствии ДНК
полимераз (A.Cornberg, M.Djulian, 1967)
получение инфекционного генетического материала фага φX174;
Химическим синтезом
(H.Corana, 1970 - 1976)- синтез тРНК аланина дрожжей (генетически неактивен);
- синтез тРНК тирозина (генетически активен), содержащий промотор (52 нуклеотида), структурный участок (126 нуклеотидов) и терминатор (21 нуклеотид);

Слайд 372. Получение рекомбинарнтных молекул ДНК
1.1. лигазный метод
Использование рестриктаз образующих “липкие
концы”
Использование лигаз
Риск нарушения целостности генов;
Не все рестриктазы образуют “липкие концы”;
Малое
количество рекомбинантных молекул;Необходимость сложного отбора рекомбинантных молекул;
1.2. терминальный метод
Использование эндо- и экзонуклеаз
Искусственное прикрепление “липких концов”
Использование в качестве вектора любых плазмид
Использование комплексных фрагментов ДНК

Слайд 383. Перенос и экспрессия рекомбинантных молекул ДНК
Типы переноса
Трансформация
В качестве
векторов используются плазмиды;
Трансфекция
В качестве векторов используются специфические фаги;
Трансгенез
В
качестве векторов используются неспецифические вирусы;Пути переноса
Прокариот → прокариот
Прокариот → эукариот
Эукариот → прокариот
Эукариот → эукариот

Слайд 39Механизмы защиты экзогенной генетической информации
Метилирование ДНК (вирусы)
Перевод линейной формы ДНК
в кольцевую (фаг λ)
Блокирование системы рестрикции клетки хозяина (фаг T3)
Действие
рестриктазПеревод линейной формы ДНК в")
Слайд 40Механизмы защиты эндогенной генетической информации
Метилирование ДНК
Действие специфических рестриктаз
Действие эригенетического окружения
Неспецифическое
действие клеточных нуклеаз
Защитные свойства мембран
Действие гистоновых белков
Действие негистоновых белков
Действие интерферона