Разделы презентаций
- Разное
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Геометрия
- Детские презентации
- Информатика
- История
- Литература
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
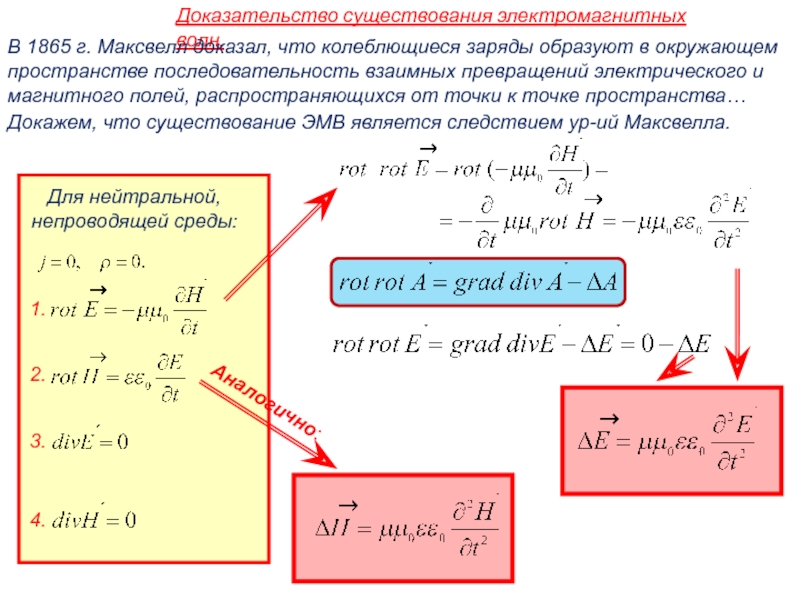
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Обмен углеводов
Содержание
- 1. Обмен углеводов
- 2. План лекции 5:Гликоген. Особенности строенияСинтез и распад
- 3. Метаболизм глюкозы
- 4. Использование метаболитов ЦТК в синтезе различных соединений
- 5. Ключевая роль глюкозо – 6-фосфата
- 6. Карта основныхпутейметаболизма
- 7. Обмен углеводов
- 8. 1. Обмен гликогенаГликогенез игликогенолиз
- 9. Гликоген – основная форма депонирования глюкозы в
- 10. В клетках гликоген находится в виде зерен.
- 11. Линейные участки молекулы гликогена образованы остатками глюкозы,
- 12. Синтез и распад гликогена Синтез и распад
- 13. Синтез гликогена - гликогенезСинтез гликогена ускоряется в
- 14. Когда длина синтезируемой цепи увеличивается на 11-12
- 15. Синтез гликогена - гликогенез
- 16. Образование УДФ-глюкозы
- 17. Далее подробно расписаны реакции гликогенеза:
- 18. 1. Получение активной формы глюкозы реакцией фосфорилирования:
- 19. 2. Изомеризация активной формы глюкозы:
- 20. 3. Образование уридиндифосфоглюкозы (УДФ-глюкозы):
- 21. Строение УДФ-глюкозыНуклеозид-фосфосахара используются как переносчики звеньев моноз в реакциях синтеза полимерных молекул
- 22. 4. Перенос остатка глюкозы с УДФ-глюкозы на
- 23. Ветвление гликогена5. Синтез α-(1,6)-гликозидных связей происходит приучастии гликоген-ветвящегося
- 24. Подводим итог:Гликоген синтезируется в период пищеварения (в
- 25. Для подготовки и включения глюкозы в растущую
- 26. Необходимость превращения глюкозы в гликоген связана с
- 27. Распад гликогена - гликогенолизили мобилизация гликогена –
- 28. Гликогенолиз -распадгликогена
- 29. 1. Освобождение глюкозы в виде глюкозо-1-фосфата (реакция
- 30. Рис. Схема гликогенолиза
- 31. Мобилизация гликогена в печени и мыщцах
- 32. Постоянный уровень глюкозы в крови в постабсорбтивный
- 33. Функция мышечного гликогена заключается в высвобождении глюкозо-6фосфата, используемого в самой мышце для окисления и получения энергии.
- 34. Рис.Особенности мобилизации гликогена в печени и мышцах
- 35. Гормоны регулирующие обмен гликогенаПереключение процессов синтеза и
- 36. Рис.Изменение активности гликогенсинтетазы и гликогенфосфорилазы
- 37. Первичным сигналом для синтеза инсулина и глюкагона
- 38. В период пищеварения преобладающим является влияние инсулина,
- 39. При интенсивной мышечной работе и стрессе в
- 40. Болезни углеводного обменаПричины: нарушения переваривания и всасывания углеводов, синтеза и распада гликогена.- ферментопатии,- патологии транспортеров моносахаридов,- гормонзависимые болезни.
- 41. Нарушения переваривания дисахаридов
- 42. Гликогеновые болезни их причины
- 43. 2. ГЛЮКОНЕОГЕНЕЗ
- 44. Карта основныхпутейметаболизма
- 45. Обмен углеводов
- 46. ГлюконеогенезГлюконеогенез – процесс синтеза глюкозы из веществнеуглеводной
- 47. Процесс протекает главным образом в печени и
- 48. Большинство реакций гликолиза и гликонеогенеза являются обратимыми
- 49. СхемаГликолиза (слева) и глюконеогенеза (справа) I, II, III – необратимые циклы,катализируютсяразными ферментами.
- 50. В митохондриях под действием ферментов малатдегидроеназы и
- 51. Превращение пирувата (ПВК) в оксалоацетат1 - транспорт
- 52. Рис.Суммарное уравнение образования глюкозы из пируватаВидно, что синтез глюкозы энергозатратный процесс
- 53. Использование лактата в качестве субстрата вглюконеогенезе связано
- 54. Цикл Кори 1 – поступление лактата из
- 55. Часть ПВК, образовавшегося из лактата, окисляется в
- 56. Снижение использования лактата в качестве субстрата в
- 57. Причиной повышения лактата в крови может быть
- 58. 3. Пентозофосфатный путь в метаболизме глюкозы
- 59. Кроме того, пентозофосфатный путь поставляет клетке пентозофосфаты
- 60. В пентозофосфатном пути превращения глюкозы можно выделить
- 61. Пентозофосфатный путь превращений глюкозы
- 62. ПФП превращения глюкозыА – окислительный этап;Б – неокислительный этап;В – пентозофосфатний цикл
- 63. Пентозофосфатный путь является альтернативным путем окисления
- 64. 3глюкозо-6-фосфат + 6 НАДФ+ →→ 3СО2 +
- 65. ПФП в эритроцитахПФП в эритроцитах очень важен,
- 66. Дефект первого фермента ПФПглюкозо-6-фосфат дегидрогеназы приводит к
- 67. Рис. Восстановление глутатиона с участием НАДФН
- 68. Благодарю за внимание
- 69. Слайд 69
- 70. Слайд 70
- 71. Общие пути катаболизмаОкисление продуктов окисления глюкозы гликолизом
- 72. Картакатаболизма
- 73. Окислительное декарбоксилирование пировиноградной кислоты Суммарный результат многостадийной реакции выглядит следующим образом:
- 74. Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в пируватдегидрогеназный комплекс (ПДК):
- 75. Этот комплекс ферментов работает подобно конвейеру, в
- 76. Дигидролипоат-ацетилтрансфераза (2) - второй фермент комплекса. Катализирует
- 77. Дегидрогеназа дигидролипоевой кислоты (3й фермент) отщепляет водород
- 78. Цикл лимонной кислотыЦитратный цикл (цикл Кребса, цикл
- 79. В ходе одного полного цитратного цикла происходит:
- 80. Цикл лимонной кислоты / цикл Кребса
- 81. Энергетика полного окисления глюкозы в клетке1. Гликолиз
- 82. Ферменты цитратного цикла 1 – цитратсинтаза, 2 – аконитаза, 3
- 83. Сопряжение общих путей катаболизма с дыхательной цепьюВ
- 84. Энергетика цитратного цикла и общих путей катаболизма
- 85. Кроме того, в цитратном цикле происходит одна
- 86. Схема взаимосвязи общего пути катаболизма и ЦПЭ
- 87. задание на дом: записать все реакции ПДК и цикла Кребса Благодарю за внимание
- 88. Использование метаболитов ЦТК в синтезе различных соединений
- 89. Метаболизм глюкозы
- 90. Схема взаимосвязи общего пути катаболизма и ЦПЭ
- 91. Синтез и распад гликогена
- 92. Синтез гликогенагликогенез
- 93. Мобилизация (распад) гликогенаПроисходит в интервалах после приема
- 94. Скачать презентанцию
План лекции 5:Гликоген. Особенности строенияСинтез и распад гликогенаГлюконеогенезПентозофосфатный путьДополнение к лекции 4 :схема «Использование метаболитов ЦТК с синтезе различных соединений»
Слайды и текст этой презентации

Слайд 2План лекции 5:
Гликоген. Особенности строения
Синтез и распад гликогена
Глюконеогенез
Пентозофосфатный путь
Дополнение к
лекции 4 :







Слайд 9Гликоген – основная форма депонирования глюкозы в клетках животных (резервный
полисахарид).
У растений эту же функцию выполняет крахмал.
В структурном
отношении гликоген, как и крахмал, представляет собой разветвленный полимер,
построенный из остатков глюкозы соединенных на линейных участках 1-4 альфа-, и в местах разветвления
1-6 альфа- гликозидными связями.
И гликоген и крахмал – гомополисахариды.
Гликоген более разветвлен и
компактен, чем крахмал.
В гликогене свыше
1 000 000 остатков глюкозы.
.")
Слайд 10В клетках гликоген находится в виде зерен. Ветвление обеспечивает быстрое
освобождение при распаде гликогена большого количества концевых мономеров - глюкозы.

Слайд 11Линейные участки молекулы гликогена образованы
остатками глюкозы, связанными
альфа-1,4-гликозидными связями,
а в местах
разветвления – альфа 1,6-гликозидными связями.
Линейные участки молекул
гликогена состоят из 6-10 остатков глюкозы.
(в крахмале линейные участки длиннее, и состоят из
20-24 остатков глюкозы). Следоватетельно, крахмал
менее разветвлен.

Слайд 12Синтез и распад гликогена
Синтез и распад гликогена гормонозависимый процессы
и регулируются инсулином и глюкагоном (и другими контринсулярными гормонами адреналином
и кортизолом).Процессы синтеза и распада гликогена не являются
обращением друг в друга, эти процессы происходят
разными путями и разными ферментами.
У животных гликоген синтезируется во всех клетках тканей органов, но особенно активно синтез идёт в мышцах и в печени, где он откладывается (в цитоплазме) в виде видных под микроскопом зёрен.
350г в мышцах,
100г в печени.

Слайд 13Синтез гликогена - гликогенез
Синтез гликогена ускоряется в период
пищеварения (абсорбтивный период
– 1-2 часа после
приёма пищи), когда глюкоза активно поступает
из крови в клетки тканей и фосфорилируется, превращаясь
в глюкозо-1- фосфат (затрачивается АТФ), а затем для
синтеза гликогена глюкозо-1-фосфат превращается в
уридин-дифосфо-глюкозу (УДФ-глюкозу).
Затрачивается УТФ, аналог АТФ
УДФ-глюкоза – субстрат для гликогенсинтетазы.
Гликогенсинтетаза переносит остаток глюкозы на
праймер (олигосахарид из 4-8 остатков глюкозы) и
соединяет его альфа-1,4- гликозидной связью.

Слайд 14Когда длина синтезируемой цепи увеличивается на
11-12 остатков глюкозы, фермент
ветвления
переносит фрагмент из 5-6 остатков глюкозы на
внутренний остаток
глюкозы , соединяя его альфа-1,6-гликозидной связью.
Удлинение цепей и ветвление их повторяется много
много (тысячи) раз. А так как в гликогене более
миллиона остатков глюкозы, то расходы энергии (АТФ и
УТФ) составляют в 2 раза большее количество молей,
чем молей глюкозы.
Т.е. синтез гликогена энергозатратный процесс.






:")
Слайд 21Строение УДФ-глюкозы
Нуклеозид-фосфосахара
используются как
переносчики
звеньев моноз
в реакциях
синтеза
полимерных
молекул

Слайд 224. Перенос остатка глюкозы с УДФ-глюкозы на затравочную цепь
гликогена, которая содержит
не менее 4-х звеньев глюкозы:
при наращивании
цепи гликогена α-(1,4)-гликозидная связьобразуется за счёт полуацетального гидроксила
добавляемого звена и гидроксильной группы в 4-м
положении остатка глюкозы растущей цепи.

Слайд 23Ветвление гликогена
5. Синтез α-(1,6)-гликозидных связей происходит при
участии гликоген-ветвящегося фермента –
гликоген-трансферазы,
которая переносит концевой
олигосахарид молекулы с невосстанавливающего
конца на гидроксильную
группу у 6-го атома углерода глюкозы той же или соседней цепи гликогена.
Благодаря способности гликогена накапливаться в
мышцах и в печени, в организме создаётся резерв
углеводов, который используется при повышении
энергозатрат организма.
-гликозидных связей происходит приучастии гликоген-ветвящегося фермента – гликоген-трансферазы, которая")
Слайд 24Подводим итог:
Гликоген синтезируется в период пищеварения
(в течение 1-2 часов
после приема углеводной пищи).
Гликогенез особенно интенсивно протекает в печени
и скелетных мышцах.
В начальных реакциях образуется УДФ-глюкоза (реакция 3),
которая является активированной формой глюкозы,
непосредственно включающейся в реакцию полимеризации
(реакция 4). Эта последняя реакция катализируется
гликогенсинтазой, которая присоединяет глюкозу к
олигосахариду или к уже имеющейся в клетке молекуле
гликогена, наращивая цепь новыми мономерами.

Слайд 25Для подготовки и включения глюкозы в растущую
полисахаридную цепь требуется
энергия
1 моль АТФ и 1 моль УTФ.
Ветвление полисахаридной
цепи происходит при участии фермента
амило- a-1,4 - a-1,6-гликозил-трансферазы
путем разрыва одной aльфа -1,4-связи и переноса
олигосахаридного остатка от конца растущей цепи
к ее середине с образованием в этом месте
aльфа -1,6-гликозидной связи .
Молекула гликогена содержит до 1 млн. остатков глюкозы,
следовательно, на синтез расходуется значительное
количество энергии.
Т.е. гликогенез энергозатратный процесс.

Слайд 26Необходимость превращения глюкозы в гликоген
связана с тем, что накопление
значительного
количества глюкозы в клетке привело бы к повышению
осмотического давления,
так как глюкоза хорошо растворимое вещество. Напротив, гликоген
содержится в клетке в виде гранул, и мало растворим.

Слайд 27Распад гликогена - гликогенолиз
или мобилизация гликогена – происходит в основном
в
период между приёмами пищи (постабсорбтивный
период) и ускоряется во время
физической работы.Этот процесс происходит путем последовательного
отщепления остатков глюкозы в виде
глюкозо-1-фосфата с помощью фермента
гликогенфосфорилазы . Этот фермент не расщепляет
альфа- 1,6- гликозидные связи в местах разветвлений,
поэтому необходимы еще 2 фермента, после действия
которых глюкозный остаток в точке ветвления
освобождается в форме свободной глюкозы. Гликоген
распадается до глюкозо-6-фосфата без затрат АТФ.


Слайд 291. Освобождение глюкозы в виде глюкозо-1-фосфата
(реакция 5) происходит в
результате фосфоролиза,
катализируемого фосфорилазой.
Фермент отщепляет концевые остатки один за
другим, укорачивая цепи гликогена. Однако этот фермент
расщепляет только a -1,4 гликозидные связи.
2. Второй фермент олигозидтрансфераза переносит
олигомерный участок ветви на линейную цепь, где
работает опять фосфорилаза.
3. Связи в точке ветвления гидролизуются ферментом
амило-a-1,6-гликозидазой , который отщепляет
мономер глюкозы в свободном виде.
 происходит в результате")

Слайд 31
Мобилизация гликогена в печени и мыщцах отличаются
В печени есть
фермент глюкозо-6-фосфорилаза,
в мышцах нет.
Этот фермент осуществляет дефосфорилирование
глюкозо-6-фосфата с образованием
свободной глюкозы, которая поступает из печени в кровь.
Следовательно главная функция гликогена печени –
резерв для поставки глюкозы в кровь в период между
приёмами пищи и использование её другими органами.
Таким образом, мобилизация гликогена печени
обеспечивает содержание глюкозы в крови на постоянном
Уровне. Это обстоятельство является обязательным
Условием для работы других органов и особенно мозга.

Слайд 32
Постоянный уровень глюкозы в крови в постабсорбтивный
период = 3,5-5,5
мМ/л
Органы мозг и эритроциты потребляют только глюкозу.
Через 10-18 часов после
приема пищи запасы гликогена значительно уменьшаются, а голодание в течение
24 часов приводит к полному его исчезновению.
Фермент глюкозо-6-фосфорилаза содержится также в
почках и клетках кишечника.
В мышцах гликоген используется только на нужды
миоцитов.

Слайд 33Функция мышечного гликогена заключается в
высвобождении глюкозо-6фосфата, используемого в
самой
мышце для окисления и получения энергии.


Слайд 35Гормоны регулирующие обмен гликогена
Переключение процессов синтеза и мобилизации
гликогена в
печени происходит при переходе состояния
пищеварения в постабсобтивный период или
состояния покоя на режим мышечной работы.
В печени – ИНСУЛИН, ГЛЮКАГОН и АДРЕНАЛИН
В мышцах – ИНСУЛИН и АДРЕНАЛИН
Действие гормонов в регулировании синтеза и распада
гликогена заключается в изменении активности 2х
ключевых ферментов:
гликогенсинтетазы и гликогенфосфорилазы путём
фосфорилировани и дефосфорилирования


Слайд 37Первичным сигналом для синтеза инсулина и
глюкагона является изменение концентрации
глюкозы в
Крови.
Инсулин и глюкагон постоянно присутствуют в крови,
при переходе
из абсорбтивного состояния в постабсорбтивное изменяется их относительная
концентрация –
инсулин/глюкагоновый индекс.
В постабсорбтивный период инсулин/глюкагоновый индекс
снижается и решающим фактором является влияние
глюкагона, который стимулирует распад гликогена в
печени.
Механизм действия глюкагона включает каскад реакций,
приводящий к активации гликогенфосфорилазы .

Слайд 38В период пищеварения преобладающим является
влияние инсулина, и инсулин/глюкагоновый индекс
в
этом случае повышается .
Инсулин способствует поступлению глюкозы в
инсулинзависимые ткани.
Под действием инсулина происходит:
А – стимуляция транспорта глюкозы в клетки мышечной ткани
Б – изменение активности ферментов путем фосфорилирования и дефосфорилирования,

Слайд 39При интенсивной мышечной работе и стрессе в кровь
из мозгового
вещества надпочечников секретируется
гормон адреналин. Он ускоряет мобилизацию
гликогена печени
и мышц, обеспечивая тем самым клетки разных тканей субстратом – глюкозой.
Действие этих гормонов в конечном счете сводится к
изменению скорости реакций катализируемых
ключевыми ферментами метаболических путей
гликогена. Активность которых регулируется
аллостерически и фосфорилированием.

Слайд 40Болезни углеводного обмена
Причины:
нарушения переваривания и всасывания углеводов,
синтеза и распада гликогена.
-
ферментопатии,
- патологии транспортеров моносахаридов,
- гормонзависимые болезни.






Слайд 46Глюконеогенез
Глюконеогенез – процесс синтеза глюкозы из веществ
неуглеводной природы.
Субстраты глюконеогенеза: пируват, лактат,
глицерол,
аминокислоты.
Важнейшей функцией глюконеогенеза является
поддержание уровня глюкозы в крови в
период длительного голодания и интенсивной физической
нагрузки.
Постоянное поступление глюкозы в клетки особенно
необходимо нервной ткани и эритроцитам.

Слайд 47Процесс протекает главным образом в печени и менее
интенсивно в корковом
веществе почек и слизистой
кишечника.
Включение различных субстратов в глюконеогенез
зависит
от физиологического состояния организма:лактат продукт анаэробного гликолиза в эритроцитах, работающих мышц и др. тканей при низком содержании О2;
глицерол продукт гидролиза жиров в жировой ткани в постабсорбтивный период и при интенсивной физической нагрузке;
аминокислоты образуются в результате распада белков мышц и соединительной ткани и включаются в глюконеогенез при длительном голодании или продолжительной мышечной нагрузке (прим.:культуризм).

Слайд 48Большинство реакций гликолиза и гликонеогенеза
являются обратимыми и катализируются одними
и теми
же ферментами.
Четыре реакции глюконеогенеза необратимы.
В целом синтез глюкозы в
глюконеогенезе можно свести к двум особенностям:
образование трехуглеродной молекулы и
конденсация двух трехуглеродных молекул в гексозу с последующим преобразованием в глюкозу.
Глюконеогенез энергозатратный процесс.

Слайд 49Схема
Гликолиза (слева) и
глюконеогенеза (справа)
I, II, III – необратимые
циклы,
катализируются
разными ферментами.
 и глюконеогенеза (справа) I, II, III – необратимые циклы,катализируютсяразными ферментами.")
Слайд 50В митохондриях под действием ферментов
малатдегидроеназы и аминотрансферазы образуется
малат
и аспартат из оксалоацетата (ЩУК), которые
пассивным антипортом удаляется из
митохондрии. В цитозоле малат и аспартат в результате
соответствующих реакций превращаются в оксалоацетат,
который декарбоксилируется и фосфорилируется под
действием фермента
Фосфоэнеол-пируват-карбокси-киназы с образованием
Фосфоэнолпирувата (ФЭП).
Все остальные реакции протекают в цитозоле.

Слайд 51Превращение пирувата (ПВК) в оксалоацетат
1 - транспорт ПВК из цитозоля
в митохондрю;
2 – превращение ПВК оксалоацетат (ОА);
3 – превращение оксалоацетата
в малат или аспартат (АСП);4 – транспорт аспартата и малата из митохондрии в цитозоль;
5 – превращение аспартата и малата в оксалоацетет в цитозоле;
6 – поступление оксалоацетата в глюконеогенез.
 в оксалоацетат1 - транспорт ПВК из цитозоля в")
Слайд 52Рис.Суммарное уравнение образования глюкозы из пирувата
Видно, что синтез глюкозы энергозатратный
процесс

Слайд 53Использование лактата в качестве субстрата в
глюконеогенезе связано с транспортом его
в печень и
превращением в ПВК.
В период интенсивного мышечного сокращении в
мышце ПВК превращается в лактат, при недостатке О2. Лактат из
мышцы кровью транспортируется в печень, где
превращается в ПВК (благодаря хорошему снабжению О2
и высокому содержанию НАД, а затем в глюкозу
(глюконеогенезом). И глюкоза опять с током крови
поступает в мышцы и эритроциты. Эту
последовательность событий назвали глюкозо-лактатным
циклом или циклом Кори.

Слайд 54Цикл Кори
1 – поступление лактата из сокращающейся мышцы и
эритроцитов с током крови в печень;
2, 3 –
синтез глюкозы в из лактата в печени;4 – поступление глюкозы из печени с током крови в работающую мышцу и в эритроциты;
5, 6 – использование глюкозы как энергетического субстрата сокращающейся мышцей и эритроцитами с образованием лактата.

Слайд 55Часть ПВК, образовавшегося из лактата, окисляется в
печени до СО2
и Н2О. Энергия выделяющаяся при
окислении, используется для синтеза АТФ,
необходимого в процессе глюконеогенеза.
Помимо печени, потребителями лактата и являются почки
и сердечная мышца, где он тоже окисляется до СО2 и Н2О
с образованием АТФ.
В мышцах в покое отношение НАД/НАДН повышается и
лактат превращается в пируват , который будет окисляться
до СО2 и Н2О с образованием АТФ.

Слайд 56Снижение использования лактата в качестве субстрата
в синтезе глюкозы, вызванное
дефектом ферментов
глюконеогенеза, и может приводить к повышению
концентрации молочной
кислоты в крови, понижением рН и, следовательно, к лактоацидозу.
Кратковременный лактоацидоз встречается довольно
часто даже у здоровых людей при интенсивной
мышечной работе, который компенсируется
гипервентиляцией легких и усиленным выведением СО2.
Н+ + НСО3- = Н2СО3 → Н2О + СО2 ↑
При некомпенсированном лактоацидозе содержание
лактата в крови увеличивается до 5 мМ/л (в норме до –
2мМ/л), значение рН крови может составлять 7,25 и менее
лактоацидоз (в норме рН крови 7,36 -7, 44)

Слайд 57Причиной повышения лактата в крови может быть нарушение метаболизма пирувата
вследствие:
тканевой гипоксии, вызывающей активацию анаэробного гликолиза;
поражений печени (токсические дистрофии, цирроз),
которые приводят к снижению утилизации лактата;наследственных дефектов ферментов глюконеогенеза;
дефектов ферментов пируватдекарбоксилазного комплекса (ПДК);
гиповитаминозов В1, В2, РР.

Слайд 583. Пентозофосфатный путь
в метаболизме глюкозы (ПФП)
Это альтернативный путь окиления
глюкозы.
К синтезу АТФ этот путь не приводит.
Этот процесс поставляет
клетке кофермент НАДФН использующийся в клетке как донор водорода в реакциях
восстановления и гидроксилирования и обеспечивает
клетки рибозо-5-фосфатом (который участвует в синтезе
нуклеотидов и нуклеиновых кислот).
Все ферменты ПФП локализованы в цитозоле клетки.
В ПФП превращения глюкозы можно выделить окислительный
и неокислительный этапы образования пентоз
Это альтернативный путь окиления глюкозы.")
Слайд 59Кроме того, пентозофосфатный путь поставляет клетке
пентозофосфаты (пентозы) ,
необходимые
для синтеза нуклеиновых
кислот и коферментов (НАД, НАДФ, ФМН и
ФАД, КоА иактивной формы витамина В12).
Все реакции пентозофосфатного пути проходят в цитозоле
клетки.
Реакции неокислительного этапа пентозофосфатного
пути являются обратимыми, поэтому становится
возможным синтез гексоз из пентоз.
Энергию клетке пентозофасфатный путь не поставляет.
 , необходимые для")
Слайд 60В пентозофосфатном пути превращения глюкозы можно
выделить две части:
А
– окислительный этап и
Б – неокислительный этап синтеза пентоз.
Окислительный этап синтеза поставляет клетке 2 основных продукта НАДФН и пентозы.
Неокислительный этап служит для образования пентоз
и может быть обратимым, следовательно, он служит и
для образования гексоз из пентоз


Слайд 62ПФП превращения
глюкозы
А – окислительный
этап;
Б – неокислительный этап;
В – пентозофосфатний
цикл

Слайд 63
Пентозофосфатный путь является
альтернативным путем окисления глюкозы.
Он включает
несколько циклов, в результате
функционирования которых из трех молекул
глюкозо-6-фосфата
образуются три молекулы СО2 и три молекулы пентоз.
Последние используются для регенерации двух
молекул глюкозо-6-фосфата и одной молекулы
глицеральдегид-3-фосфата. Поскольку из двух
молекул глицеральдегид-3-фосфата можно
регенерировать молекулу глюкозо-6-фосфата,
глюкоза может быть полностью окислена при
превращении по пентозофосфатному пути:

Слайд 643глюкозо-6-фосфат + 6 НАДФ+ →
→ 3СО2 + 2глюкозо-6-фосфат +
глицеральдегид-3-фосфат
+ 6 НАДФ-Н2
Пентозофосфатный цикл не приводит к синтезу АТФ.
Он
выполняет две главные функции:образование НАДФН для восстановительных синтезов, таких, как синтез жирных кислот и синтез стероидов (в цитоплазме клетки).
2) обеспечение рибозой синтеза нуклеотидов, синтеза нуклеиновых кислот и коферментов.

Слайд 65ПФП в эритроцитах
ПФП в эритроцитах очень важен, так как
восстановленный
НАДФН используется для защиты
клеток от активных форм кислорода.
Антиоксидант –
глутатион (ГsH) используется для превращения перекиси водорода в молекулу воды.
Затем окисленный глутатион должен восстановиться.
Восстановление окисленного глутатиона происходит с
участием восстановленного НАДФН (2 реакция).
1) Обезвреживание перекиси водорода
Н2О2 + ГsH = Н2О + Гs-sГ
2) Восстановление глутатиона:
2НАДФН + Гs-sГ = 2НАДФ + 2ГsН

Слайд 66Дефект первого фермента ПФП
глюкозо-6-фосфат дегидрогеназы приводит к
дефициту НАДФН и
невозможности восстановления
глутатиона.
Это приводит к окислению –SH групп гемоглобина
перекисью,
с последующей агрегацией протомеров гемоглобина.
Формируются тельца Хейнца.
Из за которых эритроциты теряют пластичность,
необходимую для прохождения через капилляры.
Нарушается целостность мембраны эритроцитов, что
приводит к гемолизу эритроцитов.





Слайд 71Общие пути катаболизма
Окисление продуктов окисления глюкозы гликолизом
ПВК в общих
путях катаболизма в митохондриях:
Пируват декарбоксилазным комплексом (ПДК) декарбоксилирование и дегидрирование
с образованием АцКоа.В цитратном цикле окисление АцКоА до СО2 и Н2О


Слайд 73Окислительное декарбоксилирование пировиноградной кислоты Суммарный результат многостадийной реакции выглядит следующим образом:

Слайд 74Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными
в пируватдегидрогеназный комплекс (ПДК):
:")
Слайд 75Этот комплекс ферментов работает подобно
конвейеру, в котором продукт передается
от
фермента к ферменту. Такой принцип повышает
эффективность работы ферментов, так
как снижаетслучайность в контакте реагирующих веществ с
ферментом. М.м. ПДК более 6х1000 000 Да
Далее приводятся названия ферментов и
характеристика катализируемых реакций.
Пируватдекарбоксилаза (1). В качестве кофермента в
реакции участвует тиаминдифосфат - производное
витамина В1.
Фермент катализирует отщепление карбоксильной группы
в виде СО2, а ацетильный остаток присоединяет к
липоевой кислоте - коферменту второго фермента.
Получается ацетил-липоат.

Слайд 76Дигидролипоат-ацетилтрансфераза (2) - второй фермент
комплекса. Катализирует перенос ацетильного остатка,
соединенного с липоевой кислотой на второй кофермент
HS-КоА
с образованием ацетил-КоА. Таким образом, в этой реакции участвуют два кофермента:
липоевая кислота, прочно соединенная с ферментом, и
кофермент А, объединяющийся с ферментом в момент
реакции. Водород остается связанным с липоевой кислотой,
которая превращается в дигидролипоат.
 - второй фермент комплекса. Катализирует перенос ацетильного остатка, соединенного")
Слайд 77Дегидрогеназа дигидролипоевой кислоты (3й фермент)
отщепляет водород от липоевой кислоты
и
переносит его на НАД+. Далее водород на НАДН
транспортируется в
дыхательную цепью.Главные продукты реакции - это
НАДH и ацетил-СоА.
NADH далее окисляется в дыхательной цепи,
где энергия используется на синтез 3 моль АТР, а
ацетил-СоА окисляется в цитратном цикле.
Пируватдекарбоксилазный комплекс находится на
внутренней мембране митохондрий и соединен с
ней со стороны матрикса.
 отщепляет водород от липоевой кислоты и")
Слайд 78Цикл лимонной кислоты
Цитратный цикл
(цикл Кребса, цикл трикарбоновых кислот (ЦТК)
–
это система реакций, приводящая к полному
окислению двухуглеродного ацетильного
фрагмента
до СО2 И Н2О, имеющего различное происхождение.
Цитратный цикл является общим конечным путем
окисления белков, жиров и углеводов.
Все реакции цитратного цикла,
как и окислительного декарбоксилирования
пирувата, локализованы в митохондриях.
 –это")
Слайд 79В ходе одного полного цитратного цикла происходит:
1. полное окисление
ацетильного остатка до двух молекул СО2;
2. образование трех молекул
восстановленного НАДН и одной молекулы ФAДH2; 3. образование одной молекулы ГТФ в результате субстратного фосфорилирования.
Реакции цитратного цикла,
ферменты и их характеристика приведена на
следующем слайде:


Слайд 81Энергетика полного окисления глюкозы в клетке
1. Гликолиз (распад глю до
2х ПВК) – 8 АТФ
2.
Пируват-декарбоксилазный комплекс 2 молекулы ПВК до 2 ацКоа 6 АТФ
3. 2 молекулы АцКоа до СО2 и Н2О 24 АТФ
----------------
Итого: 38 АТФ

Слайд 82Ферменты цитратного цикла
1 – цитратсинтаза,
2 – аконитаза,
3 – изоцитратдегидрогеназа,
4
– a-кетоглутаратдегидрогеназный комплекс,
5 – сукцинил-КоА-тиокиназа,
6 – сукцинатдегидрогеназа,
7 –
фумараза, 8 – малатдегидрогеназа.

Слайд 83Сопряжение общих путей катаболизма с дыхательной цепью
В общих путях катаболизма
происходит
пять реакций дегидрирования:
(одна на стадии окислительного декарбоксилирования
пирувата
и четыре в цитратном цикле). Все 10 атомов водорода переносятся на коферменты
дегидрогеназ, которые в свою очередь окисляются
в дыхательной цепи. Окисленные коферменты
возвращаются в реакции общих путей катаболизма.
Регенерация коферментов - это обязательное
условие для протекания реакции дегидрирования.
Таким образом, общий путь катаболизма
и дыхательная цепь непрерывно связаны между собой и
отдельно функционировать не могут.

Слайд 84Энергетика цитратного цикла и общих путей катаболизма
За один оборот
цитратного цикла синтезируется
12 молекул АТФ.
Девять из них образуются за
счет энергии транспорта в дыхательной цепи трех пар водорода от трех молекул НАДH + H+.Две молекулы АТР синтезируются при окислении 1 молекулы FADH2, так как в дыхательной цепи в данном случае действуют только два пункта сопряжения с окислительным фосфорилированием ADP.

Слайд 85Кроме того, в цитратном цикле происходит одна
реакция субстратного фосфорилирования,
дающая 1 моль ГТФ ( аналог АТР).
ИТОГО:
В общих путях катаболизма
синтезируется 15 молекул АТФ.
Три из них при окислительном декарбоксилировании пирувата (в ПДК) и
12 - в цитратном цикле.








Слайд 93Мобилизация (распад) гликогена
Происходит в интервалах после приема пищи
(постабсорбтивный период
или постсобтивный период)
и ускоряется после и во время физической работы.
Осуществляется
путем последовательного отщепленияостатков глюкозы, в виде глюкозо-1-фосфата ферментом
гликогенфосфорилазой с затратой молекулы Н3РО4.
В месте разветвления необходимы еще 2 фермента,
которые высвобождают глюкозу в свободном виде.
Гликоген распадается до глю-6-ф и свободной глюкозы
без затрат АТФ.
 гликогенаПроисходит в интервалах после приема пищи (постабсорбтивный период или")