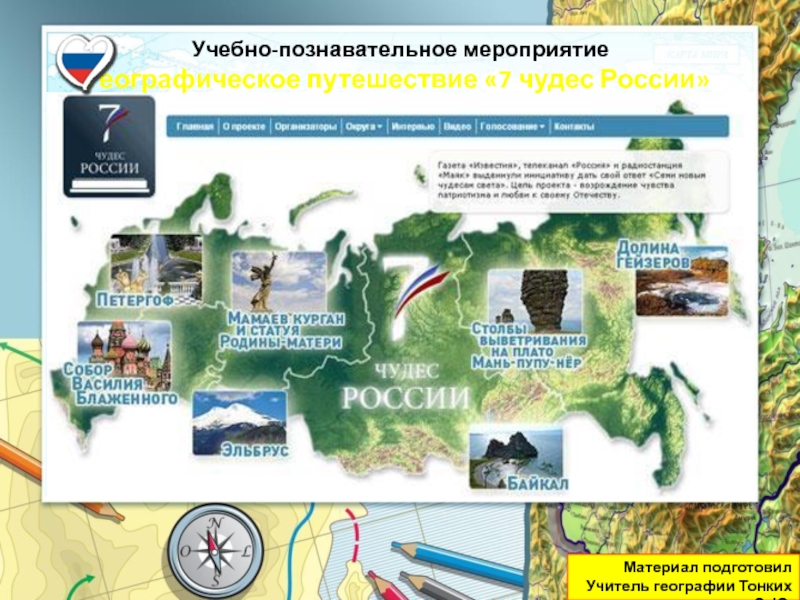
Разделы презентаций
- Разное
- Английский язык
- Астрономия
- Алгебра
- Биология
- География
- Геометрия
- Детские презентации
- Информатика
- История
- Литература
- Математика
- Медицина
- Менеджмент
- Музыка
- МХК
- Немецкий язык
- ОБЖ
- Обществознание
- Окружающий мир
- Педагогика
- Русский язык
- Технология
- Физика
- Философия
- Химия
- Шаблоны, картинки для презентаций
- Экология
- Экономика
- Юриспруденция
Тема 7. Нейрофизиология памяти. Виды памяти, процессы памяти, функциональные и
Содержание
- 1. Тема 7. Нейрофизиология памяти. Виды памяти, процессы памяти, функциональные и
- 2. Комплекс основных механизмов памяти
- 3. Этапы памяти с точки зрения выработки УР Формирование (выработка, acquisition)Консолидация (фиксация, consolidation)Хранение(retention)Воспроизведение(retrieval)
- 4. Система памятиЗа отбор информации отвечает реакция внимания — один
- 5. Система памятиСистема памяти устроена иерархично, удивительно логично
- 6. Система памятиУстановлено, что для гиппокампа характерны тета —
- 7. Виды памяти классифицируютПо форме проявления (образная, эмоциональная,
- 8. «Модальная модель» Аткинсона и Шиффрина.Понимание того, что
- 9. Поток информации в системе памяти, модель Аткинсона и Шиффрина.
- 10. Аткинсон и Шиффрин в своей трехкомпонентной модели
- 11. Если мы попытаемся охарактеризовать эти три системы
- 12. В 1972 г. Крэйк и Локхарт предложили
- 13. Сенсорная памятьЕсть, по крайней мере, два (а
- 14. Сенсорная памятьСлуховая информация дольше задерживается в сенсорной
- 15. Мгновенная (иконическая) памятьМгновенная (иконическая, сенсорная) память заключается
- 16. Кратковременная памятьЧтобы понять эту фразу, вы должны
- 17. Сравнивая функции кратковременной и долговременной памяти, можно
- 18. Кратковременная памятьРеверберационная гипотеза природы кратковременной памяти допускает
- 19. Джеймс Пейпец Круг Пейпеца гиппокамп (1)
- 20. Круг Пейпеца (а) и его упрощенная структура (б):
- 21. Круг Джеймса Пейпеца
- 22. Долговременная памятьДекларативнаясознательная, эксплицитная, о фактах и событиях.Процедурная
- 23. Ранее мною упоминалось, что идея пластичности легла
- 24. Развитие процессов стойкого облегчения проводимости сигнала На
- 25. Долгосрочная память как возникновение новых межклеточных связей
- 26. Гипотеза о роли глии в образовании временной
- 27. Итак! Механизмы памятиПревращение краткосрочной памяти в долговременную
- 28. Механизм образования кратковременной и долговременной памяти.
- 29. Нобелевская премия по физиологии и медицине 2000года
- 30. Механизмы памяти. E. Kandel (Эрик Кендел)
- 31. AplysiaМоллюску осторожно трогали сифон и тотчас вслед
- 32. За эту картинку Эрик Кендел получил Нобелевскую
- 33. Механизмы памяти. Эрик КенделИтак, Кендел обнаружил, что
- 34. Механизмы памяти. Эрик КенделБолее сильные и продолжительные
- 35. Механизмы памяти. Эрик КенделЕсли прикосновение к сифону
- 36. Механизмы памяти. Эрик Кендел.Таким образом, в отличие
- 37. ИТАК!Принцип Д. Хебба утверждает необходимость совпадения активности
- 38. PKA - Protein kinase A; PKC- Protein
- 39. Основные этапы фосфорилирования Са - и К-каналов,
- 40. Долгосрочная память как возникновение новых межклеточных связей
- 41. Сходство основных этапов процессов экспрессии генов при
- 42. Ряд препаратов, предназначенных для улучшения памяти, воздействует
- 43. Вещества, улучшающие память ( Memory enhancers)1-
- 44. В статье 2015 года Тонегава
- 45. Подобно первому исследованию 2015 года, в 2017
- 46. Тонегава разработал технику генного нокаута и показал,
- 47. Это интересно!!!! Нобелевская премия 2014 года по
- 48. Поскольку никакого подкрепления при первоначальном обследовании крысы
- 49. Нобелевская премия 2014 года по физиологии и
- 50. Нобелевская премия 2014 года по физиологии и
- 51. ДЛЯ ВАС – БЛЮДЦЕ, ДЛЯ МЕНЯ —
- 52. Биологический смысл!Итак, в кодировании всей обстановки занята
- 53. Итак, в завершение, «сухой остаток» Клеточные и
- 54. ИТАК! Человеческий мозг делится на четыре доли-
- 55. ВЫВОДЫВначале, при возникновении новой памяти гиппокамп действует
- 56. ОДНАКО!Все ныне существующие представления и гипотезы о нейрофизиологических
- 57. Уровни изучения строения и функций мозгаМолекулярныйКлеточныйСистемныйСтроениеФункции
- 58. Стратегии манипулирования нейронной активностью in vivo. Клетки
- 59. Дополнительные и обязательные для ознакомления материалы к
- 60. Механизмы памятиВозможность избирательного выключения кратковременной памяти без
- 61. Круг Джеймса Пейпеца
- 62. Механизмы памятиНа рисунке представлены механизмы участия ионов
- 63. Схема синапса, способного к развитию долговременной потенциации: 1 —
- 64. Механизмы памятиОпределенное значение в механизмах долгосрочной памяти
- 65. Итак, в итоге: молекулярный механизм формирования долговременной
- 66. Механизмы памяти. Эрик КенделНервная цепь с замкнутыми
- 67. Механизмы памятиИтак, повторю!,краткосрочная память не требует для
- 68. Механизмы памятиНесмотря на различия предположений о природе
- 69. Механизмы памятиВозможным субстратом долговременной памяти являются некоторые
- 70. Механизм образования кратковременной и долговременной памяти.
- 71. Иммунологические механизмы, повышающие проводимость определенных синапсов КБ —
- 72. Долгосрочная память как возникновение новых межклеточных связей
- 73. В дополнение: Импринтинг определен как формирование
- 74. ИмпринтингСвязи, которые развиваются между ребенком и матерью,
- 75. В дополнение: Существуют убедительные свидетельства в пользу
- 76. В дополнение: Исследователям давно известно, что питание
- 77. КАК МОЗГ СОЗДАЕТ НОВЫЕ НЕЙРОНЫ? По данным
- 78. Заключение. «Человек должен знать, что только
- 79. Механизм медленной синаптической передачи нервного импульсаПодобная универсальность
- 80. Синаптические и молекулярные механизмы памятиПоследующий анализ показал,
- 81. Механизм медленной синаптической передачи нервного импульса. Связывание
- 82. Механизмы памятиОдной из распространенных химических теорий памяти
- 83. В течение секунд и минут сохраняются изменения,
- 84. Механизм медленной синаптической передачи нервного импульсаИтак, Нобелевская
- 85. Скачать презентанцию
Комплекс основных механизмов памяти ЗапоминаниеСохранениеПоискАктивация ВоспроизведениеЗабываниепамять
Слайды и текст этой презентации
Слайд 1Тема 7. Нейрофизиология памяти. Виды памяти, процессы памяти, функциональные и
структурные основы памяти, нейрохимия памяти, нейроанатомия памяти
Людмила Николаевна
Слайд 2Комплекс основных механизмов памяти
Запоминание
Сохранение
Поиск
Активация
Воспроизведение
Забывание
память

Слайд 3Этапы памяти с точки зрения выработки УР
Формирование (выработка, acquisition)
Консолидация (фиксация,
consolidation)
Хранение(retention)
Воспроизведение(retrieval)
Консолидация (фиксация, consolidation)Хранение(retention)Воспроизведение(retrieval)")
Слайд 4Система памяти
За отбор информации отвечает реакция внимания — один из механизмов устранения
избыточности сенсорного потока и подавления многих сенсорных входов. Какую информацию мозг
хорошо запоминает? - Новую и значимую! (интересную).При нейрофизиологическом исследовании основ памяти различают собственно механизмы памяти и регуляторные механизмы памяти, к которым относятся эмоции.
Хотя поскольку память является интегральным целым (несмотря на многоуровневый характер: клеточный, молекулярный), аппарат эмоций может запускать в действие тот или иной уровень, включать деятельность тех или иных специализированных нейронов. При этом эмоциональная память включает память не только о самом состоянии эффекта, но и о системе запуска, то есть о ситуации, обусловившей его возникновение.

Слайд 5Система памяти
Система памяти устроена иерархично, удивительно логично и предусмотрительно.
Теменно — височно — затылочная
область, где замыкаются пути, идущие от различных сенсорных систем, имеет
важнейшее значение для формирования высших психических функций. Экспериментальные и клинические исследования показали, что помимо гиппокампа, в формировании памяти имеют существенное значение такие структуры мозга, как поясная извилина, передние ядра таламуса, маммилярные тела, перегородка, свод, амигдалярный комплекс, которые составляют большой и малый круг Пейпеца. Центральные фигуры — гиппокамп, ретикулярная формация, миндалина. Таким образом, мы можем нарисовать следующую картину памяти.Большой лимбический круг — гиппокампо — цингулярная (поясная извилина) система — это «информационный» цикл. Его структуры широко принимают сенсорную информацию из различных источников и последовательно обрабатывают её на разных уровнях сложности.
Второй, гиппокампо — ретикулярный цикл — регуляторный. Эта система объединяет структуры, регулирующие рабочий уровень мозга. К этому циклу примыкает дополнительная «эмоциональная» система (амигдала — гипоталамус), которая за счёт нервных и вегетативно — гормональных влияний, возникающих при эмоциях, может усиливать и продлевать возбуждение регуляторной системы.
Сигнал проходит обработку «на запись» в информационной системе только при «разрешении» со стороны регуляторной системы.

Слайд 6Система памяти
Установлено, что для гиппокампа характерны тета — волны. В процессе обучения,
создания кратковременной памяти и реакций на сигнал «что это?» в гиппокампе регистрируется
электрическая активность с частотой в пределах тета — ритма (4-7гц). Отсюда тета — ритм передаётся другим образованиям, которые настраиваются на запоминание текущего события и перенос его из «кладовой» кратковременной памяти в «кладовую» долговременной памяти.Мы можем назвать тета — ритм — ритмом внимания, начальной фазой образования условного рефлекса. Важно! Тета — ритм коррелирует с эпизодической и семантической памятью.
Как возникает гиппокампальный тета-ритм? Сенсорный сигнал достигает нейронов гиппокампа в СА1 и СА2 через энториальную кору, а затем через зубчатую фасцию и нейронов САЗ и СА4. Гиппокамп связан двусторонними связями с перегородкой (септумом). Реакция гиппокампа активирует латеральный септум и через него ретикулярную формацию (РФ), которая в свою очередь стимулирует медиальный септум, инициирующий генерацию тета-ритма. Появление гиппокампального тета-ритма связано с появлением реакций у нейронов новизны. С повторением стимула развивается привыкание за счет подключения тормозной функции зубчатой фасции, куда, так же как и в гиппокамп, приходит сенсорный сигнал из энториальной коры.

Слайд 7Виды памяти классифицируют
По форме проявления (образная, эмоциональная, логическая, или словесно-логическая)
Образная
память проявляется формированием, хранением и воспроизведением ранее воспринятого образа реального
сигнала, его нервной модели.Логическая (словесно-логическая, семантическая) память — это память на словесные сигналы, обозначающие как внешние объекты и события, так и вызванные ими ощущения и представления.
Под эмоциональной памятью понимают воспроизведение некоторого пережитого ранее эмоционального состояния при повторном предъявлении сигнала, вызвавшем первичное возникновение такого эмоционального состояния. Эмоциональная память характеризуется высокой скоростью и прочностью. Главная причина – человек легче и более устойчиво запоминает эмоционально окрашенные сигналы, раздражители. Напротив, серая, скучная информация запоминается намного труднее и быстро стирается в памяти.
По временной характеристике, или продолжительности (мгновенная (сенсорная), кратковременная, долговременная).
по характеру проявления различают механическую, условно-рефлекторную память;
по типу восприятия — зрительную, слуховую, обонятельную, двигательную и висцеральную память.
В психологии и физиологии выделяют также произвольную и непроизвольную память; Различают видовую память, обусловленную генетическими факторами, и индивидуальную, приобретаемую каждым отдельным организмом в процессе его жизни.
Образная память")
Слайд 8«Модальная модель» Аткинсона и Шиффрина.
Понимание того, что память может быть
разложена на составляющие части, не ново; оно было предложено в
1890-х годах великим американским психологом Уильямом Джеймсом, а затем Дональдом Хеббом в 1949 году. До 1960-х годов многие психологи не ощущали необходимости вводить более чем один вид памяти, но с начала 1970-х было широко воспринято некое различение между долговременной и кратковременной памятью.Активная дискуссия в 1960-х годах привела к созданию целого ряда моделей памяти упрощенного типа. Они, как правило, допускали три формы памяти — сенсорную, кратковременную и долговременную. В обобщенном виде они могут быть описаны моделью, которую предложили Ричард Аткинсон и Ричард Шиффрин (1968). Будучи типичной и при этом широко распространенной, эта модель получила название «модальная модель». Эта модель предполагает, что информация поступает из окружающей среды через параллельные ряды регистров кратковременной сенсорной памяти, а затем переходит в общее кратковременное хранилище. Считается, что эта система, действуя подобно рабочей памяти, способна оперировать информацией и переносить ее в долговременное хранилище. Действительно, кратковременное хранилище образует главное звено в этой модели; без него невозможно ни усвоение нового материала, ни воспроизведение старой информации.


Слайд 10Аткинсон и Шиффрин в своей трехкомпонентной модели добавили процесс внимания
для перевода информации из сенсорного регистра в кратковременную память для
ее дальнейшей обработки и, возможно, запоминания.А также добавили процесс структурирования, укрупнения единиц, процесс управляемого кодирования — запоминания с помощью разнообразных мнемических приемов, процессы поиска и извлечения, позволяющие вытащить информацию из долговременной памяти обратно в кратковременную, для того чтобы она могла быть использована для решения текущих задач.

Слайд 11Если мы попытаемся охарактеризовать эти три системы памяти, то про
сенсорные регистры мы можем сказать, что там информация удерживается в
полном объеме в модально-специфической форме, но не более полусекунды в зрении, не более 2–4 секунд в слухе.
Слайд 12В 1972 г. Крэйк и Локхарт предложили выделить уровни обработки
информации. Гипотеза уровневой обработки информации — гипотеза, согласно которой продолжительность
хранения информации в долговременной памяти зависит от того, каким образом данная информация была обработана.
Слайд 13Сенсорная память
Есть, по крайней мере, два (а возможно, и больше)
компонента сенсорной зрительной, или иконической, памяти, как ее иногда называют.
Один из них, по всей видимости, обусловливается сетчаткой глаза; в первую очередь на него влияет яркость предъявляемого стимула.
Второй компонент подключается в тот момент, когда мозгом получена и объединена информация с обеих сетчаток. Этот компонент иконической памяти обладает гораздо большей восприимчивостью к структуре, нежели к яркости, и представляет собой систему распознавания формы.
 компонента")
Слайд 14Сенсорная память
Слуховая информация дольше задерживается в сенсорной памяти, чем зрительная.
Это объясняет, почему последние слова в серии слов лучше запоминаются,
будучи услышанными, нежели прочитанными.Когда числовой ряд проговаривается, последний элемент ряда может быть извлечен из сенсорной слуховой памяти. Сенсорную слуховую память называют также эхоической, поскольку она имеет сходство с эхом, звучащим еще какое-то время после произнесения звукового стимула. Эхоическая память ограничена примерно одним или двумя элементами.
Последовательность произносимых цифр запоминается лучше, чем последовательность цифр, предъявляемых визуально, поскольку слуховая сенсорная память имеет более продолжительное действие.

Слайд 15Мгновенная (иконическая) память
Мгновенная (иконическая, сенсорная) память заключается в образовании мгновенного
отпечатка, следа действующего стимула в рецепторной структуре. Этот отпечаток, или
соответствующая физико-химическая энграмма внешнего стимула, отличается высокой информативностью, полнотой признаков, свойств (отсюда и название «иконическая память», т. е. четко проработанное в деталях отражение) действующего сигнала, но и высокой скоростью угасания (хранится не более 100—150 мс, если не подкрепляется, не усиливается повторным или продолжающимся стимулом).Нейрофизиологический механизм иконической памяти, очевидно, заключается в процессах рецепции действующего стимула и ближайшего последействия (когда реальный стимул уже не действует), выражаемого в следовых потенциалах, формирующихся на базе рецепторного электрического потенциала. Продолжительность и выраженность этих следовых потенциалов определяется как силой действующего стимула, так и функциональным состоянием, чувствительностью и лабильностью воспринимающих мембран рецепторных структур.
Биологическое значение иконической памяти заключается в обеспечении анализаторных структур мозга возможностью выделения отдельных признаков и свойств сенсорного сигнала, для последующего распознавания образа.
Иконическая память хранит в себе не только информацию, необходимую для четкого представления о сенсорных сигналах, поступающих в течение долей секунды, но и содержит несравненно больший объем информации, чем может быть использовано и реально используется на последующих этапах восприятия, фиксации и воспроизведения сигналов.
При достаточной силе действующего стимула иконическая память переходит в категорию краткосрочной (кратковременной) памяти.
 памятьМгновенная (иконическая, сенсорная) память заключается в образовании мгновенного отпечатка,")
Слайд 16Кратковременная память
Чтобы понять эту фразу, вы должны помнить ее начало
до тех пор, пока вы не доберетесь до ее конца.
Ее невозможно понять, если не запоминать составляющие ее слова и то, в каком порядке они расположены. Поэтому при выполнении речевых и арифметических операций имеется необходимость во временном хранилище информации для выполнения различных функций дополнительно к пониманию или вычислению. После того как задание выполнено, дополнительная информация уже больше не требуется. Этой системе, или даже скорее целому ряду систем, было дано название кратковременной, или рабочей, памяти. Там содержится информация, крайне необходимая в течение короткого периода времени, которая затем становится малозначимой. В какой степени кратковременная память отличается от долговременной?Согласно одной из точек зрения, кратковременная память представляет собой то же, что долговременная, но используется в более специальных условиях, что приводит к очень слабому долговременному сохранению информации в памяти.
Другая точка зрения состоит в том, что в долговременной и в кратковременной памяти задействованы различные системы, хотя они и очень тесно связаны в своей деятельности.
Наиболее популярна концепция временной организации памяти. принадлежащая канадскому психологу Д. Хеббу (D. Hebb), который выделил два хранилища памяти: кратковременное и долговременное.
Кратковременная память представляет собой комплексное сочетание взаимодействующих между собой подсистем, а именно рабочую память.

Слайд 17Сравнивая функции кратковременной и долговременной памяти, можно сказать, что в
кратковременной памяти мы «живем», а в долговременной памяти храним знания,
придающие смысл, значение нашему непосредственному существованию. Обращение к прошлому опыту, который необходим, чтобы понять настоящее, — это функция долговременной памяти.Кратковременная память — оперативная память, обеспечивающая выполнение текущих поведенческих и мыслительных операций. В основе кратковременной памяти лежит повторная многократная циркуляция импульсных разрядов по круговым замкнутым цепям нервных клеток. Кольцевые структуры могут быть образованы и в пределах одного и того же нейрона путем возвратных сигналов, образуемых концевыми (или боковыми, латеральными) разветвлениями аксонного отростка на дендритах этого же нейрона.
В результате многократного прохождения импульсов по этим кольцевым структурам в них постепенно образуются стойкие изменения, закладывающие основу последующего формирования долгосрочной памяти. В этих кольцевых структурах могут участвовать не только возбуждающие, но и тормозящие нейроны. Продолжительность кратковременной памяти составляет секунды, минуты после непосредственного действия соответствующего сообщения, явления, предмета.
При кратковременной памяти
Для долговременной памяти

Слайд 18Кратковременная память
Реверберационная гипотеза природы кратковременной памяти допускает наличие замкнутых кругов
циркуляции импульсного возбуждения как внутри коры большого мозга, так и
между корой и подкорковыми образованиями (в частности, таламокортикальные нервные круги), содержащими как сенсорные, так и гностические (обучаемые, распознающие) нервные клетки. Внутрикорковые и таламокортикальные реверберационные круги как структурная основа нейрофизиологического механизма краткосрочной памяти образованы корковыми пирамидными клетками V—VI слоев преимущественно лобных и теменных областей коры большого мозга.Участие структур гиппокампа и лимбической системы мозга в краткосрочной памяти связано с реализацией этими нервными образованиями функции различения новизны сигналов и считывания поступающей афферентной информации на входе бодрствующего мозга.
Реализация феномена краткосрочной памяти практически не требует и реально не связана с существенными химическими и структурными изменениями в нейронах и синапсах, так как для соответствующих изменений в синтезе матричных (информационных) РНК требуется большее время.
Несмотря на различия гипотез и теорий о природе краткосрочной памяти, исходной их предпосылкой является возникновение непродолжительных обратимых изменений физико-химических свойств мембраны, а также динамики медиаторов в синапсах. Ионные токи через мембрану в сочетании с кратковременными метаболическими сдвигами во время активации синапсов могут привести к изменению эффективности синаптической передачи, длящейся несколько секунд.

Слайд 19Джеймс Пейпец
Круг Пейпеца
гиппокамп (1) через свод (2), мамиллярные
тела (3) и передние ядра таламуса (4) связан с поясной
извилиной (5), а поясная извилина через нейроны старой коры (зубчатой извилины гиппокампа) – с гиппокампом. через свод (2), мамиллярные тела")
Слайд 20Круг Пейпеца (а) и его упрощенная структура (б): 1 — гиппокамп; 2 — свод; 3 —
мамиллярное тело; 4 — передние ядра таламуса; 5 — поясная извилина; 6 — зубчатая извилина; •
— синапсы, содержащие NMDA-рецепторыРассмотрим схему, приведенную на рис. б. На ней из всего множества структур круга Пейпеца оставлены лишь две ключевые — гиппокамп, обладающий NMDA-рецепторами, и поясная извилина, для которых характерны множественные связи с самыми разными отделами коры и подкорки (в том числе с гипоталамусом и центрами бодрствования). Гиппокамп и поясная извилина соединены множественными взаимными, формирующими круг связями (волокна свода и др.).
Как происходит фиксация информации с помощью этой системы? Представим себе человека ранним утром, его мозг полностью отдохнул. Можно считать, что при этом структуры круга Пейпеца находятся в «очищенном» состоянии, все пути передачи информации пока закрыты и магниевые пробки располагаются на своих местах — в каналах NMDA-рецепторов. Далее начинается обычный рабочий день: человек встает, умывается, завтракает, совершая обычный утренний ритуал.
Если при этом ничего необычного не происходит, то уровень возбуждения в мозге невелик и сохранение памяти идет только за счет самых нестойких суммационных механизмов. Однако, если происходит что-то необычное, значимое, имеющее эмоциональную окраску, то уровень активации в соответствующих сенсорных и ассоциативных центрах коры резко повышается. Это приводит к возбуждению поясной извилины, переадресовке сигналов на вход в гиппокамп, и, как в случае применения сильного стимула, «выбиванию магниевых пробок» в соответствующих стимулу или ситуации информационных каналах.
Момент выбивания Mg+-пробок приводит к замыканию информационного контура в круге Пейпеца, по которому затем будет происходить циркуляция возбуждения (источником слабой активации являются центры бодрствования). И пока контур функционирует — информация о прошедшем значимом событии будет периодически возвращаться через поясную извилину в кору, «напоминая» о себе.
 и его упрощенная структура (б): 1 — гиппокамп; 2 — свод; 3 — мамиллярное")
Слайд 21
Круг Джеймса Пейпеца
Из описанной схемы следует несколько выводов:
Во-первых, легко
понять, что емкость (объем памяти) круга Пейпеца хотя и велика,
но не безгранична. Каждый из нас может вспомнить ощущения, возникающие тогда, когда этот блок памяти переполняется — при просмотре нескольких кинофильмов подряд, посещении крупного музея, многочасовом сидении на лекциях.Во-вторых, система круга Пейпеца и LTP (длительная посттетаническая потенциация) оказываются чувствительными к электрошоковым воздействиям, когда тотальное выбивание магниевых пробок стирает с нейронных контуров текущую информацию. В результате человек может забыть о событиях, имевших место за несколько часов до электрошока (ретроградная амнезия).
В-третьих, восстановление ресурса этой системы, ее очистка и возврат магниевых пробок на место требует достаточно длительного прекращения циркуляции возбуждения по соответствующим контурам, что возможно лишь при выключении центров бодрствования и засыпании. При этом информация, записанная на контурах круга Пейпеца, частично теряется, а частично переходит в долговременную форму хранения.
Отметим также, что синапсы с NMDA-рецепторами встречаются не только в гиппокампе, но и в других зонах ЦНС — сенсорных центрах, двигательных центрах, ассоциативной коре больших полушарий, где они выполняют аналогичную функцию — обеспечивают быстрое формирование каналов для передачи приоритетной информации, переключают потоки сигналов с канала на канал (своеобразные «стрелочники»).

Слайд 22Долговременная память
Декларативная
сознательная,
эксплицитная, о фактах и
событиях.
Процедурная бессознательная,
имплицитная,
например, двигательные
навыки.
Декларативная:
семантическая; эпизодическая.
Семантическая –
абстрактные, безличные знания об объектах, событиях, фактах и связях между ними, никак не связанных с личным опытом.
Эпизодическая- события личной
жизни, о собственных переживаниях и
мыслях.
Образная память и когнитивные карты.
Память на пространственные ориентиры-
теория когнитивных карт. Объем когнитивной карты напрямую
влияет на процессы образного мышления и памяти.
Чем шире карта –
тем больше образов она может вместить.

Слайд 23Ранее мною упоминалось, что идея пластичности легла в основу теории,
определяющей роль обучения в формировании синаптических связей в коре больших
полушарий и объясняющей морфологические особенности обучения. Получение информации влечет за собой модуляцию синаптических контактов. Посредством суммирования совпадающих по времени входных сигналов нейроны связываются друг с другом, образуя функциональные динамические системы, причем контакты между входными и выходными нейронами усиливаются дублирующимися волокнами и обратными связями. Благодаря этим ассоциативным процессам клетки связываются в функциональные единицы памяти - «клеточные ансамбли». Основными путями реализации пластичности нервной системы и обеспечения хранения информации признаются модификация количества, конфигурации, свойств синапсов, а также изменение структуры (длина, ветвистость) отростков нейрона.
Слайд 24Развитие процессов стойкого облегчения проводимости сигнала
На рис. А показано исходное
состояние синапсов.
Если возбуждение пресинаптического нейрона ( Б) вызывает усиленное выделение
медиатора, а возбуждение постсинаптического нейрона — усиленное выделение фермента, разрушающего медиатор ( В), то при сочетаниях условного и безусловного раздражителей происходит конвергенция импульсных потоков на синапсах ( Г), которая обусловливает оптимальный баланс активности медиатора и разрушающего его фермента на стабильном высоком уровне, а также активацию сети белковых тяжей, уменьшающих просвет синаптической щели ( Д). Показано, что повышение эффективности синапсов сопровождается конформационными изменениями мембранных белков, специфичными для синаптических мембран. Кружками обозначены везикулы медиатора, треугольниками — фермент, расцепляющий медиатор.

Слайд 25Долгосрочная память как возникновение новых межклеточных связей и улучшение проведения
по уже существующим связям.
Это важно!!! Эффективность передачи возбуждения в синапсах
в значительной мере зависит от их функционального состояния, определяемого уровнем деятельности. Показано, что усиленное использование синапсов повышает их проходимость для импульсных потоков. При объяснении этого явления учитывается, что ничтожные просветы синаптических щелей соизмеримы с размерами макромолекул мембран, образующих их стенки. Поэтому многократная конвергенция импульсов условного и безусловного раздражителей на синапсах центрального нейрона, вызывая физико-химические изменения его мембран, может, в конечном итоге, сделать синаптическую щель более преодолеваемой.
Слайд 26Гипотеза о роли глии в образовании временной связи условного рефлекса.
На
стадии образования и упрочения условного рефлекса в прилегающих к нервной
клетке глиальных клетках усиливается синтез миелина, который окутывает концевые тонкие разветвления аксонного отростка и тем самым облегчает проведение по ним нервных импульсов, в результате чего повышается эффективность синаптической передачи возбуждения.Таким образом, в основе долговременной памяти могут лежать сопряженные изменения в нервно-глиальном комплексе центральных нервных образований.
А — синапс не проводит, так как потенциал возбуждения в немиелинизированной пресинаптической терминали ослабляется и медиатор выделяется в количестве, недостаточном для срабатывания синапса;
Б — глиальная клетка, возбуждаясь при безусловном подкреплении рефлекса, замыкает пресинаптический участок терминали в миелиновую оболочку и потенциал возбуждения в этой терминали сохраняет величину, необходимую для срабатывания синапса:
1 — аксон, 2 — синапс, 3 — глиальная клетка, 4 — изменение потенциала действия, 5 — миелиновая оболочка

Слайд 27Итак! Механизмы памяти
Превращение краткосрочной памяти в долговременную (консолидация памяти) в
общем виде обусловлено наступлением стойких химических и структурных изменений синаптической
проводимости как результат повторного возбуждения нервных клеток (обучающиеся популяции, ансамбли нейронов по Хеббу),По данным современной нейрофизиологии и нейрохимии, в основе долговременной (долгосрочной) памяти лежат сложные химические процессы синтеза белковых молекул в клетках головного мозга. В основе консолидации памяти много факторов, приводящих к облегчению передачи импульсов по синаптическим структурам (усиленное функционирование определенных синапсов, повышение их проводимости для адекватных импульсных потоков). Одним из таких факторов может служить феномен посттетанической потенциации, поддерживаемый реверберирующими потоками импульсов: раздражение афферентных нервных структур приводит к достаточно длительному (десятки минут) повышению проводимости нейронов.
Это означает, что возникающие при стойком сдвиге мембранного потенциала физико-химические изменения постсинаптических мембран, вероятно, служат основой для образования следов памяти, отражающихся в изменении белкового субстрата нервной клетки.
 в общем")
Слайд 28Механизм образования кратковременной и долговременной памяти. 1 — аксон; 2 — ядерная оболочка; 3
— мембрана обучающегося нейрона; 4 — рецептор к медиатору; 5 —везикулы с медиатором
Выделяющийся из
пресинаптического окончания медиатор (глутаминовая кислота) действует на соответствующие метаботропные рецепторы, запуская образование вторичного посредника (например, ц АМФ). Вторичный посредник, передавая сигнал внутри обучающегося нейрона, проникает в ядро и воздействует на ДНК. Конечным результатом такого воздействия является активация генов, управляющих синтезом рецепторов к глутаминовой кислоте: с них «снимается копия» — и-РНК, которая поступает к рибосомам. Рибосомы синтезируют новые белки-рецепторы к глутаминовой кислоте, которые встраиваются в постсинаптическую мембрану самого активного синапса, передающего информацию о характеристиках объекта. Встраивание в постсинаптическую мембрану дополнительных рецепторов является тем ключевым фактором, который приводит к увеличению эффективности синаптической передачи. Как только рецепторов окажется достаточно много, ВПСП на мембране обучающегося нейрона достигнет критического уровня, и ПД (поведенческая реакция) будет запущена. 
Слайд 29Нобелевская премия по физиологии и медицине 2000года за открытия, касающиеся
передачи сигналов в нервной системе. Paul Greengard, E. Kandel (Эрик Кендел),
Arvid Carlsson
Пол Грингард
открытие механизма действия дофамина и других нейромедиаторов
Арвид Карлссон
Исследователь дофамина

Слайд 30 Механизмы памяти. E. Kandel (Эрик Кендел) Эссе, адаптированное из его
Нобелевской речи,
озаглавлено: «молекулярная биология хранения памяти: диалог
между генами и синапсами»
Одну
из важнейших функций мозга, в которой задействованы механизмы медленной синаптической передачи и фосфорилирования белков, многие годы исследовал нобелевский лауреат, американский нейробиолог Эрик Кендел. Сначала он задался вопросом: «Почему имеются разные нейротрансмиттеры, если лишь одного достаточно для того, чтобы опосредовать передачу всех электрических сигналов?».Э.Кендел сумел показать, что для формирования самой настоящей памяти, как кратковременной, так и долговременной, достаточно всего трех нейронов определенным образом соединенных между собой. Память изучалась на гигантском моллюске, морском зайце (Aplysia). У этого моллюска относительно простая нервная система, состоящая примерно из 20 тыс. нервных клеток. Многие из них настолько велики, что видны невооруженным глазом и могут быть идентифицированы по положению в нервных ганглиях. При этом морской заяц имеет простые защитные реакции, которые можно использовать для исследования фундаментальных механизмов обучения.
 Эссе, адаптированное из его")
Слайд 31Aplysia
Моллюску осторожно трогали сифон и тотчас вслед за этим сильно
били по хвосту. После такого однократного «обучения» моллюск некоторое время
реагирует на легкое прикосновение к сифону бурной защитной реакцией, но вскоре забывает (кратковременная память). Один нейрон (сенсорный) получает сигнал от сифона, в данном случае - чувствует легкое прикосновение. Сенсорный нейрон передает импульс моторному нейрону, который, в свою очередь, заставляет сокращаться мышцы, участвующие в защитной реакции (втягивает жабру и выбрасывает в воду порцию красных чернил). Информация об ударе по хвосту поступает от третьего нейрона, который в данном случае играет роль модулирующего.
Слайд 32За эту картинку Эрик Кендел получил Нобелевскую премию в 2000
г.
На рис. показаны два синапса. Первый служит для передачи импульса
от сенсорного нейрона к моторному. Второй синапс передает импульс от модулирующего нейрона к окончанию сенсорного. Если в момент прикосновения к сифону модулирующий нейрон «молчит» (по хвосту не бьют), в синапсе 1 выбрасывается мало нейромедиатора и моторный нейрон не возбуждается. Однако удар по хвосту приводит к выбросу нейромедиатора в синапсе 2, что вызывает важные изменения в поведении синапса 1. В окончании сенсорного нейрона вырабатывается сигнальное вещество cAMP (циклический аденозинмонофосфат). Это вещество активизирует регуляторный белок протеинкиназу А. Протеинкиназа А, в свою очередь, активизирует другие белки, и это в конечном счете приводит к тому, что синапс 1 при возбуждении сенсорного нейрона (то есть в ответ на прикосновение к сифону) начинает выбрасывать больше нейромедиатора, и моторный нейрон возбуждается. Это и есть кратковременная память: пока в окончании сенсорного нейрона много активной протеинкиназы А, передача сигнала от сифона к мышцам жабры и чернильного мешка осуществляется более эффективно.
Синапс 1
Синапс 2

Слайд 33Механизмы памяти. Эрик Кендел
Итак, Кендел обнаружил, что определенные стимулы усиливают
защитный рефлекс втягивания жабры у аплизии. Эта измененная реакция сохраняется
на протяжении часов или даже дней и поэтому служит удобной моделью для изучения механизмов памяти и обучения. Исследования лаборатории Кендела показали, что в основе такой длительной реакции лежит повышение эффективности синаптической передачи между сенсорными нейронами моллюска и двигательными нервными клетками, которые активируют мышцы для защитной реакции.В основе этой формы пластичности лежит усиленный вход ионов кальция в клетку, который повышает выделение нейромедиатора сенсорным нейроном при каждом нервном импульсе и, следовательно, усиливает оборонительную реакцию. Эти изменения происходят за счет фосфорилирования белков определенных ионных каналов.

Слайд 34Механизмы памяти. Эрик Кендел
Более сильные и продолжительные стимулы формируют у
моллюска разновидности долговременной памяти, которая может длиться дни и даже
недели. Эти стимулы увеличивают содержание в клетке цАМФ и активируют протеинкиназу А. Далее такие сигналы через фосфорилирование определенных белков, так называемых транскрипционных факторов, достигают ядра нервной клетки, где меняют активность ряда генов. В результате синтез некоторых из белков заметно увеличивается, а других уменьшается. Многие из этих генов кодируют белки, участвующие в построении и функции синапсов.Благодаря каскаду молекулярных реакций изменяются функции и форма синапсов нейрона, что ведет к долговременным изменениям синаптической эффективности, лежащей в основе длительных модификаций защитного рефлекса у аплизии.

Слайд 35Механизмы памяти. Эрик Кендел
Если прикосновение к сифону сопровождается ударом по
хвосту много раз подряд, протеинкиназы А становится так много, что
она проникает в ядро сенсорного нейрона. Это приводит к активизации другого регуляторного белка — транскрипционного фактора CREB (cAMP- response element-binding protein) . Белок CREB «включает» целый ряд генов, работа которых, в конечном счете, приводит к разрастанию синапса 1 (как показано на рисунке) или к тому, что у окончания сенсорного нейрона вырастают дополнительные отростки, которые образуют новые синаптические контакты с моторным нейроном.В обоих случаях эффект один: теперь даже слабого возбуждения сенсорного нейрона оказывается достаточно, чтобы возбудить моторный нейрон. Это и есть долговременная память. Остается добавить, что, как показали дальнейшие исследования, у высших животных и у людей память основана на тех же принципах, что и у аплизии.

Слайд 36Механизмы памяти. Эрик Кендел.
Таким образом, в отличие от кратковременной памяти,
требующей фосфорилирования уже присутствующих в клетке белков, долговременная память основывается
на экспрессии генов и синтезе новых белков.Значит, если заблокировать синтез белков в нервной системе, исчезает долговременная память, а кратковременная остается неповрежденной?
Замечательная особенность этой цепи клеточных процессов состоит в том, что фундаментальные ее звенья и компоненты чрезвычайно схожи при обучении у моллюсков и у млекопитающих, оставаясь, по-видимому, неизменными на протяжении многих миллионов лет эволюции нервной системы.
Это позволило Кенделу начиная с 90-х годов перенести значительную часть обнаруженных им на моллюске закономерностей на модели сложного обучения у мышей. Используя технологию гомологических рекомбинаций, позволяющую удалять у этих животных отдельные гены, Кендел и его сотрудники показали, что основные компоненты молекулярного каскада формирования памяти, описанные им для аплизии, необходимы и при консолидации памяти у млекопитающих.

Слайд 37ИТАК!
Принцип Д. Хебба утверждает необходимость совпадения активности пресинаптического и постсинаптического
нейронов как условие усиления связи между ними (пре-постсинаптическое совпадение). Он
получил название «синапс Хебба».Согласно второму принципу (Э.Кендел )для усиления связи между нейронами активность постсинаптической клетки не нужна, если одновременно с пресинаптической клеткой активирован третий нейрон — модулирующий (премодулирующее совпадение).
Суть обучения, зависящего от активности, составляет одновременная активация фермента аденилатциклазы ионами Са, входящими в нейрон во время генерации ПД, и серотонином от модулирующих нейронов.

Слайд 38PKA - Protein kinase A; PKC- Protein kinase C; IP3-
inositol trisphosphate; PLC - Фосфолипаза C; CS- conditioned (условный) stimulus
CS ; US- unconditioned (безусловный) stimulus
Слайд 39Основные этапы фосфорилирования Са - и К-каналов, обеспечивающего кратковременное изменение
проводимости синапса. Особая функция ионов кальция в передаче сигнала объясняется
тем, что он в пресинаптическом нейроне запускает цепочку биохимических процессов, которая увеличивает число активированных Са-каналов. Это создает эффект усиления выброса медиатора из пресинаптического окончания.Последовательность биохимических событий, развертывающихся в пресинаптическом нейроне после входа в него ионов Са, выглядит следующим образом. В нейроне для ионов Са существует внутренний рецептор— кальмодулин. Это специфический белок, с которым кальций образует комплекс. Возникнув, последний активирует фермент аденилатциклазу. С ее помощью из АТФ синтезируется циклический аденозинмонофосфат (цАМФ), активирующий протеинкиназы. Протеинкиназа А фосфорилирует белки потенциал-независимых кальциевых каналов в результате присоединения к ним фосфатной группы. Это открывает дополнительное число Са-каналов. В клетку входит еще большее количество ионов кальция, что дополнительно увеличивает выход медиатора из нейрона. Ионы кальция, как и цАМФ, выполняют роль посредников в выбросе медиатора из пресинаптической мембраны. Работа клеточного аппарата, обеспечивающего преобразование ПД в выход медиатора и тем самым передачу сигнала через химический синапс от нейрона к нейрону, представлена на рисунке (вход I).Этот же аппарат клетки, обеспечивающий передачу сигнала от нейрона к нейрону, используется и для длительных изменений синаптической проводимости, связанных с обучением: привыканием, сенситизацией, а также с ассоциативным обучением, главным образом на начальном его этапе. Критическая роль в этих процессах принадлежит модулирующим нейронам. Они имеют свой вход к этому клеточному аппарату — вход II. У моллюсков это серотонинергические нейроны. Внутриклеточной мишенью для серотонина, так же как для комплекса кальций+кальмодулин, является аденилатциклаза, за счет которой из аденозинтрифосфата (АТФ) синтезируется циклический аденозинмонофосфат (цАМФ). За счет двойного действия на аденилатциклазу Са и серотонина возрастает образование цАМФ. Активированные киназы фосфорилируют белки ионных каналов. Киназа А взаимодействует с белками кальциевых каналов и открывает их для входа ионов кальция. Киназа С фосфорилирует белки калиевых каналов, вызывая их закрытие.

Слайд 40Долгосрочная память как возникновение новых межклеточных связей и улучшение проведения
по уже существующим связям.
Не
вызывает сомнений, что в основе всякого обучения лежит процесс проторения путей возбуждения. Наглядной иллюстрацией того, что в результате обучения потоки нервных импульсов легче и быстрее преодолевают синаптическую задержку, может служить модельный опыт с тараканом: после ампутации передних лапок, которыми он чистит усики, таракан обучается пользоваться для этой цели задними лапками. При этом время синаптической задержки моторных импульсов к задним лапкам резко уменьшается. Наконец, зависимость количества синапсов (по показателям электронной микроскопии) от интенсивности функционирования нервных механизмов продемонстрирована в экспериментах с повышением активности синапсов путем предъявления сложных задач или снижения активности синапсов путем сенсорной депривации.
По другим предположениям долгосрочная память реализуется облегчением передачи импульсов по существующим синапсам. Это может быть достигнуто стойкими изменениями структуры и свойств синапсов.

Слайд 41Сходство основных этапов процессов экспрессии генов при развитии и обучении.
Экстраклеточные сигналы активируют вторичные мессенджеры, которые высвобождают каталитические субъединицы протеинкиназ.
Протеинкиназа транспортируется в ядро клетки и там через транскрипционные факторы действует на ранние гены (c-fos и c-jun), вызывая их экспрессию. Последние кодируют транскрипционные факторы для поздних генов — белки: FOS и JUN. Эти белки и вызывают экспрессию поздних генов, которая определяет синтез различных структурных белков и молекул, необходимых для синаптического роста.Э. Кэндел- теория каскада молекулярных реакций при обучении предполагает три уровня памяти. Кратковременная память, которая длится от нескольких минут до нескольких часов, обусловлена ковалентной модификацией белков, не достигших состояния возбуждения. Промежуточная память, охватывающая несколько часов, обусловлена фосфорилированием белков. Долговременная память, длящаяся более чем один день, зависит уже от индукции новых генов через вторичных посредников и регуляторов транскрипции. Э. Кэндел предположил, что ранние эффекторные гены ответственны за синтез белков, которые сохраняют память в течение дней. Память, сохраняемая в течение недель и месяцев, поддерживается другими — поздними эффекторными генами, которые включаются ранними регуляторными генами.

Слайд 42Ряд препаратов, предназначенных для улучшения памяти, воздействует главным образом на
два процесса, развивающихся в нейронах во время консолидации памяти:
1. Деполяризацию
мембраны и 2. Активацию СRЕВ белка.
Напомню! Деполяризация возникает после того, как высвобождение возбуждающего нейротрансмиттера глутамата в синапс стимулирует АМРА рецепторы на поверхности нейрона, получающего нервный сигнал. Под влиянием деполяризации на глутамат реагирует и другой поверхностный белок — NMDA рецептор. В результате внутри клетки активируется сложная последовательность молекулярных взаимодействий, включающая образование циклического АМФ и, как следствие, активацию СRЕВ белка. (Прерывистыми стрелками обозначены звенья этого процесса, опущенные для упрощения схемы.) Последнее событие имеет решающее значение для консолидации памяти: активированный СRЕВ помогает «включить» гены, ответственные за синтез белков, укрепляющих определенные синапсы.
Некоторые другие препараты ускоряют процессы памяти за счет усиления реакции АМРА рецепторов на глутамат, т.е. благодаря облегчению деполяризации.
А третьи увеличивают уровень активного СRЕВ в клетках — например, за счет подавления активности фермента фосфодиэстеразы, которая разрушает циклический АМФ.
11
МЕМ 1414 и некоторые другие препараты пролонгируют активацию СRЕВ пути, подавляя активность фосфодиэстеразы
Препараты, получившие название ампакинов, усиливают реакцию АМРА рецепторов на глутамат
AMPA рец.
NMDA рец.
1
2
3
4
5
ядро
ген
Активированный CREB
фосфодиэстераза
Циклический AMP
1.- Активация AMPA рецептора приводит к деполяризации мембраны
2.- Активация NMDA рецептора после деполяризации мембраны вызывает образование циклического AMP
3.- Циклический AMP вызывает активацию CREB
4.- CREB способствует активации определенных генов
5.- Активированные гены начинают синтез белков, увеличивающих эффективность синапсов
Белки,повыш.эффективн.
Аксонная терминаль нейрона,посылающего сигнал
глутамат

Слайд 43Вещества, улучшающие память
( Memory enhancers)
1- Rolipram-ингибитор фосфодиэстеразы-4, активатор cAMP-
CREB.
2-Donepezil, Amiridin-ингибиторы холинэстеразы.
3-D-cyclozerine-частичный агонист NMDA рецепторов.
4-Ampakines-позитивные модуляторы AMPA
рецепторов. Увеличивают длительность открытого состояния AMPA - каналов.1- Rolipram-ингибитор фосфодиэстеразы-4, активатор cAMP- CREB.2-Donepezil,")
Слайд 44
В статье 2015 года Тонегава и его коллеги впервые показали,
что воспоминания могут сохраняться даже тогда, когда блокируется синтез клеточных
белков. Они обнаружили, что воспоминания можно искусственно извлечь с использованием оптогенетической техники. Исследователи назвали эти ячейки памяти «тихими энграммами».«Тихие энграммы» также могут сформироваться и при различных нейродегенеративных заболеваниях типа болезни Альцгеймера. Учёные доказали: воспоминания остаются, только к ним теряется доступ, который в принципе можно восстановить (на мышиной модели болезни Альцгеймера).
На более позднем этапе изучения системной консолидацией памяти, исследователи обнаружили энграммы которые кодировали одну и ту же память как в гиппокампе, так и в префронтальной коре.
Однако соответствующие участки префронтальной коры «молчали» примерно две недели после того, как память первоначально записалась, но при этом участки гиппокампа активировались сразу. Со временем всё становилось наоборот, и замолкали участки гиппокампа.
Фото на конфокальном микроскопе. Зелёным окрашены энграммные клетки гиппокампа CA1 со встроенным в их мембрану светочувствительным белком channelrhodopsin-2. Синим выделены все остальные клетки, расположенные в области дорсального гиппокампа.
Если заблокировать синтез белков в нервной системе, исчезает долговременная память, а кратковременная остается неповрежденной?
Сусуму Тонегава (Susumu Tonegawa) лауреат Нобелевской премии по физиологии и медицине 1987 года «За открытие генетического принципа образования разнообразия антител». В последнее время переключился на исследование клеточных механизмов образования памяти и подтвердил теорию об энграммных клетках, в которых записывается память.

Слайд 45Подобно первому исследованию 2015 года, в 2017 году они обучали
мышей помнить, в какой клетке они испытывали шоковую реакцию.
Сразу после
тренировки ингибировался синтез клеточных белков, и мыши таким образом забывали о полученном опыте. Но когда под воздействием света клетки памяти активировались (а всем животным предварительно в мембраны нейронов встраивался светочувствительный белок), мыши замирали в страхе даже в нейтральном месте. Оказалось, что эти воспоминания можно восстанавливать на протяжении восьми дней после первоначального обучения.Этими результатами поддержалась гипотеза Тонегавы о том, что усиление синаптических соединений, необходимое для первоначальной кодировки памяти, не является необходимым для последующего длительного хранения. Вместо этого он говорит, что память хранится в конкретном шаблоне соединений, образованных между ансамблями энграммных клеток. Эти соединения, которые очень быстро формируются при кодировании, отличаются от синаптического усиления, которое происходит позже (в течение нескольких часов после события) с помощью синтеза необходимых белков.
Это поставило вопрос о том, зачем тогда после кодирования синтезируются белки. Учитывая, что «тихие энграммы» не извлекаются естественным путём, исследователи полагают, что основная цель белкового производства – обеспечить возможность естественной реакции на вызовы, чтобы память работала максимально эффективно.

Слайд 46Тонегава разработал технику генного нокаута и показал, что нокаут одной
субъединицы (R1) в NMDA рецепторе нейрона поля СА1 приводит к
потере долговременной памяти и ухудшению ориентации в пространстве.
Слайд 47Это интересно!!!! Нобелевская премия 2014 года по физиологии и медицине-
Джону О’Кифу и супругам Мозер, Эдварду и Мэй-Бритт за «открытия
клеток, которые образуют систему навигации в мозге».
С самого начала этих исследований было предположено, что обнаруженные свойства нейронов гиппокампа соответствуют представлению о «когнитивных картах» Эдварда Толмена, который показал, что крысы, имевшие возможность обследовать лабиринт, лучше обучаются последующим навыкам в этом лабиринте, чем крысы, которые такой возможности не имели.

Слайд 48
Поскольку никакого подкрепления при первоначальном обследовании крысы не получали, Толмен
предположил, что такие «когнитивные карты» формируются как абстрактные представления, а
не как цепочка подкрепляемых моторных реакций.Кроме нейронов, активирующихся при определенной ориентации животного, позднее были описаны нейроны именно «места» (place cells), активирующиеся в определенном месте вне зависимости от конкретной ориентации животного в пространстве; нейроны «направления головы» (head direction cells); нейроны «координатной сетки» (grid cells); нейроны «границ» (border cells). Не так давно были описаны нейроны, маркирующие трехмерное пространство у летучих мышей.

Слайд 49Нобелевская премия 2014 года по физиологии и медицине- Джону О’Кифу
и супругам Мозер, Эдварду и Мэй-Бритт за «открытия клеток, которые
образуют систему навигации в мозге».Каждый нейрон «координатной сетки» активируется в нескольких местах обследуемого пространства, так что карта активаций выглядит как гексагональная решетка, где каждый узел решетки — активация. А нейроны «границ» активируются там, где границы предоставляемого к обследованию пространства заканчиваются.
Описанные нейроны можно было бы представить компонентами системы навигации мозга, некоего «внутреннего» навигатора, сообщающего нам, где мы.
Однако есть несколько моментов, которые усложняют эту картину. Судя по тому, что последовательность активаций нейронов места у животных повторяется в моменты остановок и во сне, а также у людей при припоминании эпизода перемещения в виртуальном пространстве, наша память о пространстве может воспроизводиться в виде комбинаций активаций определенных групп нейронов. В недавних исследованиях были описаны так называемые клетки «времени» гиппокампа.

Слайд 50Нобелевская премия 2014 года по физиологии и медицине- Джону О’Кифу
и супругам Мозер, Эдварду и Мэй-Бритт за «открытия клеток, которые
образуют систему навигации в мозге».
Можно предположить, что нейронные группы гиппокампа представляют собой опыт организма, специальным образом организованный в пространстве и времени. А учитывая, что определенная последовательность активаций клеток гиппокампа может возникать еще до того, как животное попало в данное пространство , это должно быть скорее не навигацией, а предсказаниями мозга относительно будущих событий, основанными на опыте прошлого.

Слайд 51ДЛЯ ВАС – БЛЮДЦЕ, ДЛЯ МЕНЯ — ГНЕЗДО. Мышь наслаждается покоем
в блюдце, которое принимает за гнездо.
В ходе экспериментов было случайно
обнаружено, что небольшое количество нейронов гиппокампа реагировало на абстрактное представление о «гнезде». Эти клетки интенсивно отвечали на гнезда любых типов независимо от
того, были ли они круглыми, квадратными или треугольными, сделанными из хлопка, пластмассы или дерева.
То есть, некоторые нейронные клетки в гиппокампе кодируют абстрактные представления. Оказалось, что отдельные нейроны мышей реагируют на предметы различной формы и текстуры лишь при условии, что в них имеется доступное углубление, позволяющее использовать их в качестве гнезда. Закройте углубление, и клетки перестанут реагировать.

Слайд 52Биологический смысл!
Итак, в кодировании всей обстановки занята целая популяция клеток.
Одни клетки кодируют направление движения животного, другие - клетки «места».
Эти две области вместе координируют движения и направления движения животного. Данный факт особенно важен для выживания в дикой природе, когда животное после поиска пищи должно возвратиться в свою нору, гнездо, и тд. Эта навигационная система обучения связана с семантической и эпизодической памятью. У животных возникает устойчивое представление о расположении предметов на их территории – аналогично нашему фактическому обучению об окружающем нас мире. И в этой карте помещаются данные об опасности – где был замечен хищник в последний раз. Клетки места фиксируют больше, чем просто место, они помогают животным запоминать, где произошли события.Присужденная Нобелевская премия важна тем, что отмечает достижения науки в области понимания того, что лежит в основе «высших», когнитивных функций. Это демонстрирует связь активности нейронов с субъективными абстрактными «концепциями», такими как «место», и в очередной раз показывает, что мы можем иметь доступ к «внутреннему» миру организма. Существенно и то, что акцент изучения «высших» функций мозга смещается от поиска структур, осуществляющих те или иные функции, к поиску распределенных по мозгу нейронных групп, активирующихся в соответствии с той или иной решаемой организмом задачей.

Слайд 53Итак, в завершение, «сухой остаток»
Клеточные и молекулярные события при
консолидации памяти обычно развиваются в течение первых минут или часов
после обучения и приводят к возникновению перестроек в нейронах (нервных клетках) или группах нейронов. Позднее на протяжении дней или даже лет может происходить гораздо более медленная консолидация на системном уровне, при которой реорганизуются нервные сети, занятые обработкой множества отдельных воспоминаний. Процесс консолидации, влияющий на декларативную память, т.е. на воспоминания об отвлеченных фактах и конкретных событиях, опирается на работу гиппокампа и других медиальных височных структур мозга. На клеточном уровне память выражается в изменениях структуры и функций нейронов. Например, могут возникать новые синапсы , что позволяет строить новые нейронные сети.В качестве альтернативы могут усиливаться уже существующие синапсы, обеспечивая нейронам большую восприимчивость при обмене информацией друг с другом. Консолидация этих синаптических изменений требует синтеза новой РНК и белков в гиппокампе, что позволяет превратить временные модификации синаптической передачи в устойчивые перестройки синаптической архитектуры. С течением времени изменяются и вышележащие мозговые структуры.

Слайд 54ИТАК! Человеческий мозг делится на четыре доли- лобную, височную, теменную,
затылочную доли.
Лобная доля связана с более высокими когнитивными функциями, такими
как исполнительное суждение и планирование, а также вознаграждение, мотивация, внимание и определенная кратковременная память. Височная доля связана со зрительной и слуховой памятью и речью.
Теменная доля обрабатывает соматосенсорную информацию.
Затылочная доля включает зрительную кору, где визуальная информация обрабатывается и распространяется.
Теперь известно, что медиальная височная доля, часть височной доли мозга, участвует в формировании, закреплении и кратковременном хранении новых воспоминаний, особенно декларативных воспоминаний.
Медиальная височная доля структурно состоит из гиппокампа, периферического, энторинального отделов, и парагиппокампальной коры, и миндалины.
Гиппокамп является ключевым регионом для формирования и консолидации о воспоминаниях

Слайд 55ВЫВОДЫ
Вначале, при возникновении новой памяти гиппокамп действует совместно с сенсорными
областями новой коры (самого наружного слоя головного мозга). Элементы памяти
о любом событии нашей жизни распределены по многочисленным областям коры в соответствии с их содержанием. Например, зрительная информация обрабатывается в первичной зрительной коре (в затылочной доле на заднем полюсе мозга), в то время как звуковая информация анализируется первичной слуховой корой (в височных долях по бокам мозга).Когда память только формируется, гиппокамп сразу же объединяет разрозненные фрагменты информации в единое воспоминание, выступая в качестве каталога отдельных записей, хранящихся в различных сенсорных областях коры. С течением времени благодаря ряду клеточных и молекулярных процессов происходит усиление прямых связей между областями новой коры, что позволяет осуществлять доступ к памяти уже без помощи гиппокампа. Таким образом, в то время как поражение гиппокампа при травме или нейродегенеративном заболевании (например, болезни Альцгеймера) нарушает возможность образования новой декларативной памяти,память о фактах и событиях, уже успевшая пройти консолидацию на системном уровне, может сохраниться.

Слайд 56ОДНАКО!
Все ныне существующие представления и гипотезы о нейрофизиологических основах памяти не являются
до конца изученными и доказанными . В этой связи и на сегодняшний день
эта проблема интригующе интересна как для физиологов, так и для психологов.

Слайд 58Стратегии манипулирования нейронной активностью in vivo. Клетки нервной системы находятся
в гетерогенных популяциях, состоящих из различных подтипов (здесь изображены в
виде различных цветов). Методы электрической микростимуляции имеют высокую временную точность, но влияют на все клетки и волокна, окружающие электрод. Физические зонды необратимо аблируют или обратимо охлаждают клетки, чтобы уменьшить нервную активность, но также воздействуют на все клетки и волокна, окружающие электрод. Фармакологическая инъекция лекарственного средства может быть нацелена на определенный тип клеток на основе клеточной экспрессии белка, но лекарственное средство может оставаться в системе в течение нескольких минут или часов после инъекции. Генетическая инактивация определенных клеток (таких как клетки, изображенные синим цветом) также не имеет временной точности и часто необратима. Оптогенетическая стимуляция или ингибирование клеток с помощью света позволяет осуществлять специфическое нацеливание оптогенетических зондов клеточного типа (таких как клетки, изображенные синим цветом) с точностью до миллисекунды времени активации (TRENDS in Molecular Medicine)Electrical
Physical
Pharmacological
Genetic
Optogenetic

Слайд 59Дополнительные и обязательные для ознакомления материалы к теме.
Виды памяти
В результате
процесса обучения возникают физические, химические и морфологические изменения в нервных
структурах, которые сохраняются некоторое время и оказывают существенное влияние на деятельность организма. Совокупность таких структурно-функциональных изменений в нервных образованиях, становится важным фактором, определяющим все разнообразие приспособительного адаптивного поведения организма. Такие структурно-функциональные изменения известны под названием «энграмма» (след) действующих раздражителей.Память-способность к воспроизведению прошлого опыта, одно из основных свойств нервной системы, выражающееся в способности длительно хранить информацию о событиях внешнего мира и реакциях организма и многократно вводить её в сферу сознания и поведения.

Слайд 60Механизмы памяти
Возможность избирательного выключения кратковременной памяти без нарушения долговременной и
избирательного воздействия на долговременную память в отсутствие каких-либо нарушений краткосрочной
памяти обычно рассматривается как свидетельство разной природы лежащих в их основе нейрофизиологических механизмов.Косвенным доказательством наличия определенных различий в механизмах кратковременной и долговременной памяти являются особенности расстройств памяти при повреждении структур мозга.
Так, при некоторых очаговых поражениях мозга (поражения височных зон коры, структур гиппокампа), при сотрясении мозга, наступают расстройства памяти, выражающиеся в потере способности запоминать текущие события или события недавнего прошлого (произошедшие незадолго до воздействия, вызвавшего данную патологию) при сохранении памяти на прежние, давно случившиеся события. Однако ряд других воздействий оказывает однотипное влияние и на кратковременную, и на долговременную память.
По-видимому, несмотря на некоторые заметные различия физиологических и биохимических механизмов, ответственных за формирование и проявление кратковременной и долговременной памяти, в их природе намного больше общего, чем различного; их можно рассматривать как последовательные этапы единого механизма фиксации и упрочения следовых процессов, протекающих в нервных структурах под влиянием повторяющихся или постоянно действующих сигналов.

Слайд 61
Круг Джеймса Пейпеца
Устойчивая активация глутаматных синапсов – основной способ формирования
долговременной памяти, значительная часть которой – ассоциации между любыми сенсорными
стимулами и любыми реакциями (двигательными и вегетативными).Эти ассоциации – основа поведенческих программ, значимость которых растет при успешной реализации (на нейронном уровне такой рост означает все более выраженную активацию синаптической передачи).
Новые каналы для передачи информации формируются между нейронами коры за счет повышения эффективности соответствующих синапсов. В основе: активация деятельности главного возбуждающего медиатора мозга – глутаминовой кислоты (глутамата).

Слайд 62Механизмы памяти
На рисунке представлены механизмы участия ионов кальция в долговременной
потенциации – блокирование калиевых каналов, и в долговременной памяти –
активация синтеза РНК, синтез протеинкиназ и изменение количества мембранных рецепторов.
Слайд 63Схема синапса, способного к развитию долговременной потенциации: 1 — метаботропный (не-NMDA-peцептор); 2 — NMDA-рецептор,
находящийся в нерабочем состоянии; 3 — везикула с глутаматом; 4 — хемочувствительный ионный канал
; 2 — NMDA-рецептор, находящийся")
Слайд 64Механизмы памяти
Определенное значение в механизмах долгосрочной памяти имеют и изменения,
наблюдающиеся в медиаторных механизмах, обеспечивающих процесс химической передачи возбуждения с
одной нервной клетки на другую. В основе пластических химических изменений в синаптических структурах лежит взаимодействие медиаторов, например ацетилхолина с рецепторными белками постсинаптической мембраны и ионами (Na+, K+, Са2+).Динамика трансмембранных токов этих ионов делает мембрану более чувствительной к действию медиаторов. Установлено, что процесс обучения сопровождается повышением активности фермента холинэстеразы, разрушающей ацетилхолин, а вещества, подавляющие действие холинэстеразы, вызывают существенные нарушения памяти.

Слайд 65Итак, в итоге: молекулярный механизм формирования долговременной памяти. При изучении защитного
рефлекса (втягивание жабры в ответ на стимуляцию сифона) у морского
зайца установлено, что нейромедиаторы, действующие на клетку, вызывают в ней каскад молекулярных реакций (показано стрелками). В результате изменяется форма и функции синапсов, что приводит к длительным модификациям защитного рефлекса.
Слайд 66Механизмы памяти. Эрик Кендел
Нервная цепь с замкнутыми нейронными цепями, по
которым могут циркулировать потоки импульсов (по Э. Кенделу):
1 — корковый нейрон,
получающий афферентные сигналы, 1А и 1Б — промежуточные нейроны, 2 — эфферентный нейрон; белыми треугольниками изображены возбуждающие синапсы, черными — тормозящий, стрелками показано направление движения потоков импульсов.(краткосрочная память)

Слайд 67Механизмы памяти
Итак, повторю!,краткосрочная память не требует для своей реализации синтеза
информационных (матричных) молекул РНК и ДНК. Она и не могла
бы обеспечиваться этими биохимическими процессами как генетическая память, поскольку включение в полимер ДНК или РНК нуклеотидов происходит в сотни и тысячи раз медленнее, чем это необходимо для запечатления в них полной информации о текущих событиях. Вместе с тем, при любом понимании ее природы, в том числе и реверберационном, речь идет о протекании потоков импульсов по новым для них путям, т.е. о временной синаптической проводимости.
Слайд 68Механизмы памяти
Несмотря на различия предположений о природе краткосрочной памяти, их
объединяет рассмотрение механизма памяти как возникновения непродолжительных обратимых физико-химических свойств мембран
и динамики медиаторов в синапсах, временно перестраивающих поведение нервных сетей. Эти изменения неизбежно связаны со сложной цепочкой далеко идущих, сравнительно более стойких перестроек метаболизма нервной клетки.Поэтому кратковременная память может оставлять более длительный скрытый след, на основе которого, в некоторых случаях, она способна восстанавливаться после действия амнестического агента.
Углубление и структурно-метаболическое закрепление этих следов, по-видимому, является механизмом перехода краткосрочной памяти в долгосрочную.
В отличие от краткосрочной, долгосрочная память хранит следы пережитых событий неопределенно долго. Собственно она и является носителем жизненного опыта животного; знаний, приобретенных человеком.

Слайд 69Механизмы памяти
Возможным субстратом долговременной памяти являются некоторые пептиды гормональной природы,
простые белковые вещества, специфический белок S-100. К таким пептидам, стимулирующим,
например, условно-рефлекторный механизм обучения, относятся некоторые гормоны (АКТГ, соматотропный гормон, вазопрессин и др.).Гипотеза И. П. Ашмарина основана на признании важной роли активной иммунной реакции в консолидации, формировании долгосрочной памяти. Суть этого представления состоит в следующем: в результате метаболических процессов на синаптических мембранах при реверберации возбуждения на стадии формирования кратковременной памяти образуются вещества, играющие роль антигена для антител, вырабатываемых в глиальных клетках. Связывание антитела с антигеном происходит при участии стимуляторов образования медиаторов или ингибитора ферментов, разрушающих эти стимулирующие вещества.

Слайд 70Механизм образования кратковременной и долговременной памяти. 1 — аксон; 2 — ядерная оболочка; 3
— мембрана обучающегося нейрона; 4 — рецептор к медиатору; 5 —везикулы с медиатором
Выделяющийся из
пресинаптического окончания медиатор (глутаминовая кислота) действует на соответствующие метаботропные рецепторы, запуская образование вторичного посредника (например, ц АМФ). Вторичный посредник, передавая сигнал внутри обучающегося нейрона, проникает в ядро и воздействует на ДНК. Конечным результатом такого воздействия является активация генов, управляющих синтезом рецепторов к глутаминовой кислоте: с них «снимается копия» — и-РНК, которая поступает к рибосомам. Рибосомы синтезируют новые белки-рецепторы к глутаминовой кислоте, которые встраиваются в постсинаптическую мембрану самого активного синапса, передающего информацию о характеристиках объекта. Встраивание в постсинаптическую мембрану дополнительных рецепторов является тем ключевым фактором, который приводит к увеличению эффективности синаптической передачи. Как только рецепторов окажется достаточно много, ВПСП на мембране обучающегося нейрона достигнет критического уровня, и ПД (поведенческая реакция) будет запущена. 
Слайд 71Иммунологические механизмы, повышающие проводимость определенных синапсов
КБ — катионные белки, открывающие
путь комплексам фрагментов антигена синапса с РНК через оболочку клетки;
Л — клетки глии, функционально подобные лимфоцитам, М — клетки глии, функционально подобные макрофагам; 1, 2, 3 — фрагменты синаптических антигенов (зоны их синтеза заштрихованы), 4 — комплекс фрагмента антигена с РНК, 5 — антитела для синапсов, тонкими линиями изображена мРНК, толстыми — ДНК хромосом
Слайд 72Долгосрочная память как возникновение новых межклеточных связей и улучшение проведения
по уже существующим связям.
Физико-химические изменения мембран, возникающие при стойком сдвиге их
поляризации, могут служить основой для формирования памятных следов. Такие сдвиги поляризации воспроизведены в экспериментах с созданием зон повышенной возбудимости в коре при помощи постоянного тока и, видимо, имеют место в естественных условиях возникновения доминантных очагов. Опытным путем показано, что создаваемое в доминантном очаге коры состояние возбудимости подпорогового уровня под действием приходящего афферентного возбуждения может достичь порога проявления ответной реакции. Образуется суммационный рефлекс, который при закреплении следов проторения новых путей становится настоящим условным рефлексом.
Характерная особенность такого рефлекса состоит в том, что по показателям нейронной активности он протекает в две фазы: торможения и последующей отдачи (возбуждения). Образование таких временных связей происходит преимущественно в нейронах нижних слоев коры.

Слайд 73 В дополнение: Импринтинг определен как формирование в некоторые периоды онтогенеза устойчивой
индивидуальной избирательности по отношению к внешним стимулам.
Принципиальная схема механизма, связанного
с формированием импринтинга: 1 — обучающийся синапс; 2 — обучающийся нейрон; 3 — двигательный нейрон Схема механизма импринтинга на молекулярном уровне: 1 — аксон; 2 — ядерная оболочка; 3 — мембрана обучающегося нейрона; 4 — рецептор к медиатору; 5 —везикулы с медиатором

Слайд 74Импринтинг
Связи, которые развиваются между ребенком и матерью, были изучены на
примере молодых цыплят в процессе, называемом импринтингом. Теперь мы знаем,
где происходит процесс обучения в головном мозге цыпленка и какие химические трансмиттеры высвобождаются для фиксации «изображения» матери. Это изображение настолько точное, что цыпленок всегда следует исключительно за своей матерью. Молодые животные также нуждаются в понимании того, какая пища безопасна, и посредством собственного опыта учатся различать плохой вкус от хорошего. Этот факт не может быть обусловлен только генетической предрасположенностью – происходит также обучение. При импринтинге или поедании пищи активируются соответствующие рецепторы, приводящие к выработке каскада вторичных химических мессенджеров, которые приводят к активации соответствующих генов и выработке соответствующих белков в клетках мозга, дословно удерживающих память.
Слайд 75В дополнение:
Существуют убедительные свидетельства в пользу того, что дисфункция NMDA
рецепторов способна объяснить патофизиологию позитивных, негативных и когнитивных симптомов шизофрении.
NMDA рецепторы разбросаны по всему мозгу, их работа критична для процессов обучения и запоминания, развития мозга и общей нейронной обработки информации. Исходя из этого, нарушение работы NMDA рецепторов может оказаться ключевым фактором в процессе, который некоторые исследователи прозвали «болезнь синапса». Помимо других функций, рецептор регулирует выброс дофамина в других нейронных цепях; следовательно, NMDA-гипотеза не отрицает, а дополняет и развивает дофаминовую теорию.Нормальная работа NMDA рецептора полагается на три ключевых фактора: связывание глутамата с рецепторным сайтом, деполяризация содержащей рецептор нейронной мембраны и наличие глицина. Это вещество является незаменимым вторичным фактором для работы рецепторов NMDA; в его отсутствие рецептор не работает правильно. Многие предлагаемые сейчас методы улучшения работы NMDA рецепторов основаны на применении глицина. Первые свидетельства о роли NMDA рецепторов в шизофрении появились почти так же случайно, как и свидетельства для дофаминовой гипотезы; их дали наблюдения за людьми, принимавшими химический состав, который изменяет нормальную сигнализацию рецепторов NMDA. Когда здоровые добровольцы получали дозу кетамина (вещество, блокирующее активность рецептора NMDA), они демонстрировали широкий спектр распространенных симптомов шизофрении . Объекты исследования воспроизводили не только психотические симптомы, но и спектр негативных симптомов, у них были отмечены нейрокогнитивные нарушения, нарушение плавного отслеживания глазами движущихся предметов, и нарушения нейронных потенциалов (последние два признака отмечались у больных шизофренией в ходе различных исследований). Последующие исследования показали, что страдающие шизофренией люди, вероятно, имеют повышенный уровень натурального антагониста NMDA-рецепторов (аббревиатура этого вещества - NAAG). Повышенный уровень вещества отмечен в предлобной коре, височной коре, и гиппокампе . По результатам исследований, низкий уровень глицина может являться ключевым фактором в нарушении активности NMDA рецепторов при шизофрении. Выявленные ответственные гены действуют по-разному, но их объединенное влияние приводит к началу центрального патофизиологического процесса: изменению синаптической пластичности, особенно отражающемуся на глутаматной передаче сигналов через NMDA рецепторы.

Слайд 76В дополнение:
Исследователям давно известно, что питание ночью отрицательно сказывается на
здоровье организма, и теперь они решили проверить, как это отражается
на работе мозга. Для того, чтобы это выяснить, нейробиолог Кристофер Колвелл вместе с коллегами из Калифорнийского университета использовали лабораторных мышей, которые жили по привычному для них режиму, то есть были активными ночью и спали в течение дня. Некоторых животных из этой группы ученые начали кормить днем, то есть в то время, когда они должны спать.Мыши, которых кормили в неположенное время суток, быстро привыкли просыпаться днем, чтобы поесть. Время их сна сместилось, однако в сумме они спали столько же, сколько и их собратья с нормальным распорядком дня, а также ели и весили столько же, сколько и мыши из другой группы.
Затем ученые исследовали память мышей. В одном эксперименте они поместили два различных предмета в коробки с мышами, чтобы те изучили новые объекты. А спустя некоторое время ученые снова посадили животных в клетки с двумя предметами, однако теперь один из объектов был уже знаком животным, а другой – нет.
Авторы исследования наблюдали за тем, как долго мыши обнюхивали предметы. Так, по результатам эксперимента оказалось, что у мышей со сдвинутым распорядком дня сильно ухудшилась память. Эти животные провели значительно больше времени, исследуя уже знакомый объект, будто бы они не помнили, что обнюхивали этот предмет раньше.
Во втором эксперименте ученые решили напугать животных, а спустя определенное время снова поместили их в то же место, где животные почувствовали страх. Как оказалось, мыши с неправильным режимом питания испытывали меньший страх, поскольку, вероятно, не помнили о том, что произошло с ними в том месте.
Наконец, прием пищи в неправильное время суток сказался и на обучаемости мышей. Ученые обнаружили, что мыши, которые питались в неположенное время суток, медленнее учились, нежели особи из другой группы.
Ученые точно не знают, будет ли прием еды в неправильное время суток, оказывать такое же воздействие на когнитивные возможности людей, но данные исследования, безусловно, вызывают озабоченность по этому поводу. Так что если у вас ухудшилась память, возможно, вам просто стоит отказаться от поздних ужинов.

Слайд 77КАК МОЗГ СОЗДАЕТ НОВЫЕ НЕЙРОНЫ?
По данным некоторых ученых, нейрональные
стволовые клетки дают начало новым клеткам мозга.
Они периодически делятся
в двух основных областях: в желудочках (фиолетовый цвет),которые заполнены спинномозговой жидкостью, питающей центральную нервную систему, и в гиппокампе (голубой цвет) — структуре, необходимой для обучения и памяти. При пролиферации стволовых клеток (внизу) образуются новые стволовые клетки и клетки предшественники, которые могут превратиться либо в нейроны, либо в поддерживающие клетки, называемые глиальными (астроциты и дендроциты).
Однако дифференцировка новорожденных нервных клеток может произойти только после того, как они уйдут прочь от своих предков, что удается в среднем лишь половине из них, а остальные гибнут. Во взрослом мозге новые нейроны были обнаружены в гиппокампе и обонятельных луковицах, необходимых для восприятия запахов.
Ученые надеются заставить взрослый мозг восстанавливаться, вызывая деление и развитие нейрональных стволовых клеток или клеток-предшественников там и тогда, где и когда это необходимо.
Глиальные клетки

Слайд 78Заключение. «Человек должен знать, что только и только посредством мозга
возникают наши удовольствия, смех и шутки, точно, как также скорбь, боль,
горе и страхи. Посредством мозга мы думаем, видим, слышим и можем отличить прекрасное от уродливого, плохое от хорошего, приятное от неприятного» Гиппократ – 5 век до н.э.Изучение мозга оказалось невозможно без привлечения различных наук. В английском языке для обозначения науки о мозге используется слово — «neuroscience», которое дословно переводится как «нейронаука». Это отражает тот факт, что исследованием мозга занимаются не только биологи и медики, но и, например, физики, математики. Системный подход, с одной стороны, помогает лучше решить поставленную задачу, с другой, является источником разногласий.
Специалисты в различных областях говорят на разных научных языках и порой с трудом понимают друг друга. Здесь будет к месту вспомнить восточную сказку о трех слепцах, которые трогали слона, и пытались понять, на что тот похож. Один прикоснулся к хоботу и утверждал, что слон похож на шланг. Тот, что взялся за хвост, предполагал, что он похож на шнурок. Третий, думал, что слон похож на колонну, поскольку он обхватил руками его ногу.
Так же происходит в нейронауке. Можно выделить три основных подхода к исследованию мозга. Во-первых — «сверху вниз» (энцефалография, компьютерная томография). В этом случае мозг рассматривается как целостная система, состоящая из связанных динамически подключающихся структур. Второй подход — «снизу вверх» (исследования на молекулярно-клеточном уровне). Ученые пытаются понять, как работает клеточная молекулярная машина, позволяющая клеткам мозга выполнять их функции и объединяться в локальные сети. Третий подход — «мозг — черный ящик». Он осуществляется тогда, когда фармакологи пытаются непосредственно связать свойства того или иного рецептора с поведением. Нейрогенетики хотят найти ген, отвечающий за ту или иную функцию мозга. Такой подход не сильно информативен в плане понимания механизмов работы мозга, но позволяет добиваться практически важных результатов (например, в создании лекарств или диагностике генетических заболеваний).
Что же лучше? Скальпель хирурга, физиологический электрод, биохимический анализ или компьютерная томография? Вероятно, все методы хороши, если они позволяют накапливать знания для последующих обобщений.

Слайд 79Механизм медленной синаптической передачи нервного импульса
Подобная универсальность роли медленной синаптической
передачи в формировании памяти, безусловно, открывает новые возможности биохимической коррекции
нарушенной памяти у человека. Действительно, несколько лет назад Кендел основал биотехнологическую компанию, направленную на поиск принципиально новых средств регуляции памяти. Эти исследования особенно важны потому, что болезнь Альцгеймера и другие виды возрастных патологий (особенно характерных для развитых стран) начинаются именно с нарушений памяти.Все ныне существующие представления и гипотезы о нейрофизиологических основах памяти не являются до конца изученными и доказанными . В этой связи и на сегодняшний день эта проблема интригующе интересна как для физиологов, так и для психологов.

Слайд 80Синаптические и молекулярные механизмы памяти
Последующий анализ показал, что ДП обусловлена
активностью рецепторов по обеим сторонам синапса. Когда пресинаптический нейрон получает
высокочастотную стимуляцию, то выделяется нейротрансмиттер глутамат. При этом для осуществления ДП постсинаптический, или получающий, нейрон должен быть приведен в рабочее состояние на таком же частотном уровне. Соответствующие постсинаптические рецепторы обладают чувствительностью к веществу, известному под аббревиатурой NMDA , и разряд зависит от наличия точного баланса ионов в рецепторном канале. Когда пре- и постсинаптические условия оказываются правильными, природа синапса изменяется таким образом, что в будущем более слабый пресинаптический стимул будет вызывать разряд постсинаптического нейрона. Известно, что клетки, связанные с ДП, особенно многочисленны в гиппокампе, который решающим образом задействован в процессах обучения и памяти, и это укрепляет нашу убежденность в том, что этот механизм и в самом деле может быть основным механизмом обучения. В классической работе, опубликованной в 1949 году, канадский психолог Дональд Хебб предполагает, что механизм, подобный этому, может лежать в основе процессов обучения. С тех пор было разработано множество компьютеризированных моделей обучения, использующих идеи Хебба.
Слайд 81Механизм медленной синаптической передачи нервного импульса. Связывание дофамина с рецепторами
на клеточной мембране повышает в клетке содержание «вторичного посредника» -
цАМФ. Это активирует фермент протеинкиназу, которая фосфорилирует мембранные белки ионных каналов, благодаря чему осуществляется регуляция передачи нервных импульсов.Итак, Кендал показал, что кратковременная память обусловлена фосфорилированием белков, образующих в клеточных мембранах ионные каналы, через которые могут проходить ионы кальция и другие ионы, участвующие в передаче нервного импульса. А долговременную память обеспечивает синтез новых белков, запускаемый в результате воздействия сильных стимулов. Эти белки изменяют форму синапса и его чувствительность к последующим стимулам. Так была разрешена загадка, остававшаяся нерешенной в течение многих лет: почему для долговременной памяти необходим синтез белков, а для кратковременной – нет?

Слайд 82Механизмы памяти
Одной из распространенных химических теорий памяти была гипотеза Хидена
о белковой природе памяти.
По мнению автора, информация, лежащая в
основе долговременной памяти, кодируется, записывается в структуре полинуклеотидной цепи молекулы. Разная структура импульсных потенциалов, в которых закодирована определенная сенсорная информация в афферентных нервных проводниках, приводит к разной перестройке молекулы РНК, к специфическим для каждого сигнала перемещениям нуклеотидов в их цепи. Таким образом происходит фиксация каждого сигнала в виде специфического отпечатка в структуре молекулы РНК. Исходя из гипотезы Хидена, можно предположить, что глиальные клетки, принимающие участие в трофическом обеспечении функций нейрона, включаются в метаболический цикл кодирования поступающих сигналов путем изменения нуклеотидного состава синтезирующих РНК. Весь набор вероятных перестановок и комбинаций нуклеотидных элементов обеспечивает возможность фиксировать в структуре молекулы РНК огромный объем информации.
Итак, процесс фиксации информации в нервной клетке находит отражение в синтезе белка, в молекулу которого вводится соответствующий следовой отпечаток изменений в молекуле РНК. При этом молекула белка становится чувствительной к специфическому узору импульсного потока, тем самым она как бы узнает тот афферентный сигнал, который закодирован в этом импульсном паттерне. В результате происходит освобождение медиатора в соответствующем синапсе, приводящее к передаче информации с одной нервной клетки на другую в системе нейронов, ответственных за фиксацию, хранение и воспроизведение информации.

Слайд 83В течение секунд и минут сохраняются изменения, вызываемые стимулом в
NMDA-рецепторах, отвечающих на глутамат, а также в мускариновых рецепторах, чувствительных
к АХ. Сохранение следового эффекта в течение минут или часов определяется действием киназ, которые фосфорилируют пресинаптические белки ионных каналов, а также последующей экспрессией ранних генов (c-fos, c-jun). Время сохранения памяти от 1 до 6 ч связывают с синтезом новых белков — глюкопротеинов в результате экспрессии поздних генов. Глюкопротеины создают структурные изменения, встраиваясь в пресинаптические и постсинаптические участки мембран. Период 12-24 ч связан с образованием дополнительных шипиков, увеличением их диаметра и площади постсинаптических уплотнений, которые выявляются с помощью электронного микроскопа. Постсинаптические уплотнения образованы плотно пригнанными рецепторами.
Слайд 84Механизм медленной синаптической передачи нервного импульса
Итак, Нобелевская премия 2000 г.
за исследование механизмов «медленной синаптической передачи», достойно завершила историю изучения
клеточных основ деятельности мозга в ХХ в. Что же дальше? Кендел еще в 1963 г., после известия о награждении Нобелевской премией А.Ходжкина, А.Хаксли и Дж.Эклса за изучение мембранных процессов нервного возбуждения и торможения, заявил, что следующая премия будет присуждена за исследование синаптических механизмов памяти. И взялся за изучение этого вопроса. Теперь понятно, что, хотя его труд по достоинству увенчался наградой, о которой он мечтал, Кендел ошибся как минимум дважды. Как это не раз бывало с присуждением Нобелевских премий, он получил ее за исследование не той проблемы, которой посвятил всю свою жизнь. Более того, за столько лет, истекших с момента его предсказания, около дюжины Нобелевских премий присуждено за исследования мозга и ни одна из них - за расшифровку механизмов памяти. Современная нейронаука слишком мало знает о механизмах высших функций мозга, и на долю следующего века остается еще много фундаментальных открытий, касающихся этого самого сложного из всех известных нам во Вселенной объектов.